we are increasingly assailed with science-related “news” – stories that too often involve hype and attempts to garner attention (and no, half-baked ideas are not theories, they are often non-scientific speculation or unconstrained fantasies).
The other day, as is my addiction, I turned to the “Real Clear Science” website to look for novel science-based stories (distractions from the more horrifying news of the day). I discovered two links that seduced me into clicking: “Atheism is not as rare or as rational as you think” by Will Gervais and Peter Sjöstedt-H’s “Consciousness and higher spatial dimensions“. A few days later I encountered “Consciousness Is the Collapse of the Wave Function” by Stuart Hameroff. On reading them (more below), I faced the realization that science itself, and its distorted popularization by both institutional PR departments and increasingly by scientists and science writers, may be partially responsible for the absurdification of public discourse on scientific topics [1]. In part the problem arises from the assumption that science is capable of “explaining” much more than is actually the case. This insight is neither new nor novel. Timothy Caulfield’s essay Pseudoscience and COVID-19 — we’ve had enough already focuses on the fact that various, presumably objective data-based, medical institutions have encouraged the public’s thirst for easy cures for serious, and often incurable diseases. As an example, “If a respected institution, such as the Cleveland Clinic in Ohio, offers reiki — a science-free practice that involves using your hands, without even touching the patient, to balance the “vital life force energy that flows through all living things” — is it any surprise that some people will think that the technique could boost their immune systems and make them less susceptible to the virus?” That public figures and trusted institutions provide platforms for such silliness [see Did Columbia University cut ties with Dr. Oz?] means that there is little to distinguish data-based treatments from faith- and magical-thinking based placebos. The ideal of disinterested science, while tempered by common human frailties, is further eroded by the lure of profit and/or hope of enhanced public / professional status and notoriety. As noted by Pennock‘ “Science never guarantees absolute truth, but it aims to seek better ways to assess empirical claims and to attain higher degrees of certainty and trust in scientific conclusions“. Most importantly, “Science is a set of rules that keep the scientists from lying to each other. [2]
It should surprise no one that the failure to explicitly recognize the limits, and evolving nature of scientific knowledge, opens the door to self-interested hucksterism at both individual and institutional levels. Just consider the number of complementary/alternative non-scientific “medical” programs run by prestigious institutions. The proliferation of pundits, speaking outside of their areas of established expertise, and often beyond what is scientifically knowable (e.g. historical events such as the origin of life or the challenges of living in the multiverse which are, by their very nature, unobservable) speaks to the increasingly unconstrained growth of pathological, bogus, and corrupted science which, while certainly not new [3], has been facilitated by the proliferation of public, no-barrier, no-critical feedback platforms [1,4]. Ignoring the real limits of scientific knowledge and rejecting, or ignoring, the expertise of established authorities, rejects the ideals that have led to science that “works”.
Of course, we cannot blame the distortion of science for every wacky idea; crazy, conspiratorial and magical thinking may well be linked to the cognitive “features” (or are they bugs) of the human brain. Norman Cohn describes the depressing, and repeated pattern behind the construction of dehumanizing libels used to justify murderous behaviors towards certain groups [5]. Recent studies indicate that brains, whether complex or simple neural networks, appear to construct emergent models of the world, models they use to coordinate internal perceptions with external realities [6]. My own (out of my area of expertise) guess is that the complexity of the human brain is associated with, and leads to the emergence of internal “working models” that attempt to make sense of what is happening to us, in part to answer questions such as why the good die young and the wicked go unpunished. It seems likely that our social nature (and our increasing social isolation) influences these models, models that are “checked” or “validated” against our experiences.
It was in this context that Gervais’s essay on atheism caught my attention. He approaches two questions: “how Homo sapiens — and Homo sapiens alone — came to be a religious species” and “how disbelief in gods can exist within an otherwise religious species?” But is Homo sapiens really a religious species and what exactly is a religion? Is it a tool that binds social groups of organisms together, a way of coping with, and giving meaning to, the (apparent) capriciousness of existence and experience, both, or something else again? And how are we to know what is going on inside other brains, including the brains of chimps, whales, or cephalopods? In this light I was struck by an essay by Sofia Deleniv “The ‘me’ illusion: How your brain conjures up your sense of self” that considers the number of species that appear to be able to recognize themselves in a mirror. Turns out, this is not nearly as short a list as was previously thought, and it seems likely that self-consciousness, the ability to recognize yourself as you, may be a feature of many such systems. Do other organisms possess emergent “belief systems” that help process incoming and internal signals, including their own neural noise? When the author says, “We then subtly gauge participants’ intuitions” by using “a clever experiment to see how people mentally represent atheists” one is left to wonder whether there are direct and objective measures of “intuitions” or “mental representations”? Then the shocker, after publishing a paper claiming that “Analytic Thinking Promotes Religious Disbelief“, the authors state that “the experiments in our initial Science paper were fatally flawed, the results no more than false positives.’ One is left to wonder did the questions asked make sense in the first place. While it initially seemed scientific (after all it was accepted and published in a premiere scientific journal), was it ever really science?
Both “Consciousness and Higher Spatial Dimensions” and “Consciousness Is the Collapse of the Wave Function”, sound very scientific. Some physicists (the most sciencey of scientists, right?) have been speculating via “string theory” and “multiverses”, a series of unverified (and likely unverifiable) speculations, that they universe we inhabit has many many more than the three spatial dimensions we experience. But how consciousness, an emergent property of biological (cellular) networks, is related to speculative physics is not clear, no matter what Nobel laureates in physics may say. Should we, the people, take these remarks seriously? After all these are the same folks who question the reality of time (for no good reason, as far as I can tell, as I watch my new grandchild and myself grow older rather than younger).
Part of the issue involves what has been called “the hard problem of consciousness”, but as far as I can tell, consciousness is not a hard problem, but a process that emerges from systems of neural cells, interacting with one another and their environment in complex ways, not unlike the underlying processes of embryonic development, in which a new macroscopic organism composed of thousands to billions of cells emerges from a single cell. And if the brain and body are generating signals (thoughts) then in makes sense these in turn feed back into the system, and as consciousness becomes increasingly complex, these thoughts need to be “understood” by the system that produced them. The system may be forced to make sense of itself (perhaps that is how religions and other explanatory beliefs come into being, settling the brain so that it can cope with the material world, whether a nematode worm, an internet pundit, a QAnon wack-o, a religious fanatic, or a simple citizen, trying to make sense of things.
Thanks to Melanie Cooper for editorial advice and Steve Pollock for checking my understanding of physics; all remaining errors are mine alone!
Scheufele, D. A. and Krause, N. M. (2019). Science audiences, misinformation, and fake news. Proceedings of the National Academy of Sciences116, 7662-7669
Kenneth S. Norris, cited in False Prophet by Alexander Kohn (and cited by John Grant in Corrupted Science.
See Langmuir, I. (1953, recovered and published in 1989). “Pathological science.” Research-Technology Management 32: 11-17; “Corrupted Science: Fraud, Ideology, and Politics in Science” and “Bogus Science: or, Some people really believe these things” by John Grant (2007 and 2009)
21st Century DEVO-2 In the first post in this series [link], I introduced the observation that single celled organisms can change their behaviors, often in response to social signals.They can respond to changing environments and can differentiate from one cellular state to the another. Differentiation involves changes in which sets of genes are expressed, which polypeptides and proteins are made [previous post], where the proteins end up within the cell, and which behaviors are displayed by the organism. Differentiation enables individuals to adapt to hostile conditions and to exploit various opportunities.
The ability of individuals to cooperate with one another, through processes such as quorum sensing, enables them to tune their responses so that they are appropriate and useful. Social interactions also makes it possible for them to produce behaviors that would be difficult or impossible for isolated individuals.Once individual organisms learn, evolutionarily, how to cooperate, new opportunities and challenges (cheaters) emerge. There are strategies that can enable an organism to adapt to a wider range of environments, or to become highly specialized to a specific environment,through the production of increasingly complex behaviors.As described previously, many of these cooperative strategies can be adopted by single celled organisms, but others require a level of multicellularity.Multicellularity can be transient – a pragmatic response to specific conditions, or it can be (if we ignore the short time that gametes exist as single cells) permanent, allowing the organism to develop the range of specialized cells types needed to build large, macroscopic organisms with complex and coordinated behaviors. In appears that various forms of multicellularity have arisen independently in a range of lineages (Bonner, 1998; Knoll, 2011). We can divide multicellularity into two distinct types, aggregative and clonal – which we will discuss in turn (1). Aggregative (transient) multicellularity:Once organisms had developed quorum sensing, they can monitor the density of related organisms in their environment and turn or (or off) specific genes (or sets of genes, necessary to produce a specific behavior.While there are many variants, one model for sucha behavior isa genetic toggle switch, in which a particular gene (or genes) can be switched on or off in response to environmental signals acting as allosteric regulators of transcription factor proteins (see Gardner et al., 2000).Here is an example of an activity (↓) that we will consider in class to assess our understanding of the molecular processes involved.
One outcome of such a signaling system is to provoke the directional migration of amoeba and their aggregation to form the transient multicellular “slug”.Such behaviors has been observedin a range of normally unicellular organisms (see Hillmann et al., 2018)(↓). The classic example isthe cellular slime mold Dictyostelium discoideum (Loomis, 2014).Under normal conditions, these
unicellular amoeboid eukaryotes migrate, eating bacteria and such. In this state, the range of an individual’s movement is restricted to short distances.However when conditions turn hostile, specifically a lack of necessary nitrogen compounds, there is a compelling reason to abandon one environment and migrate to another, more distant that a single-celled organism could reach. This is a behavior that depends upon the presence of a sufficient density (cells/unit volume) of cells that enables them to: 1) recognize one another’s presence (through quorum sensing), 2) find each other through directed (chemotactic) migration, and 3) form a multicellular slug that can go on to differentiate. Upon differentiation about 20% of the cells differentiate (and die), forming a stalk that lifts the other ~80% of the cells into the air.These non-stalk cells (the survivors) differentiate into spore (resistant to drying out) cells that are released into the air where they can be carried to new locations, establishing new populations.
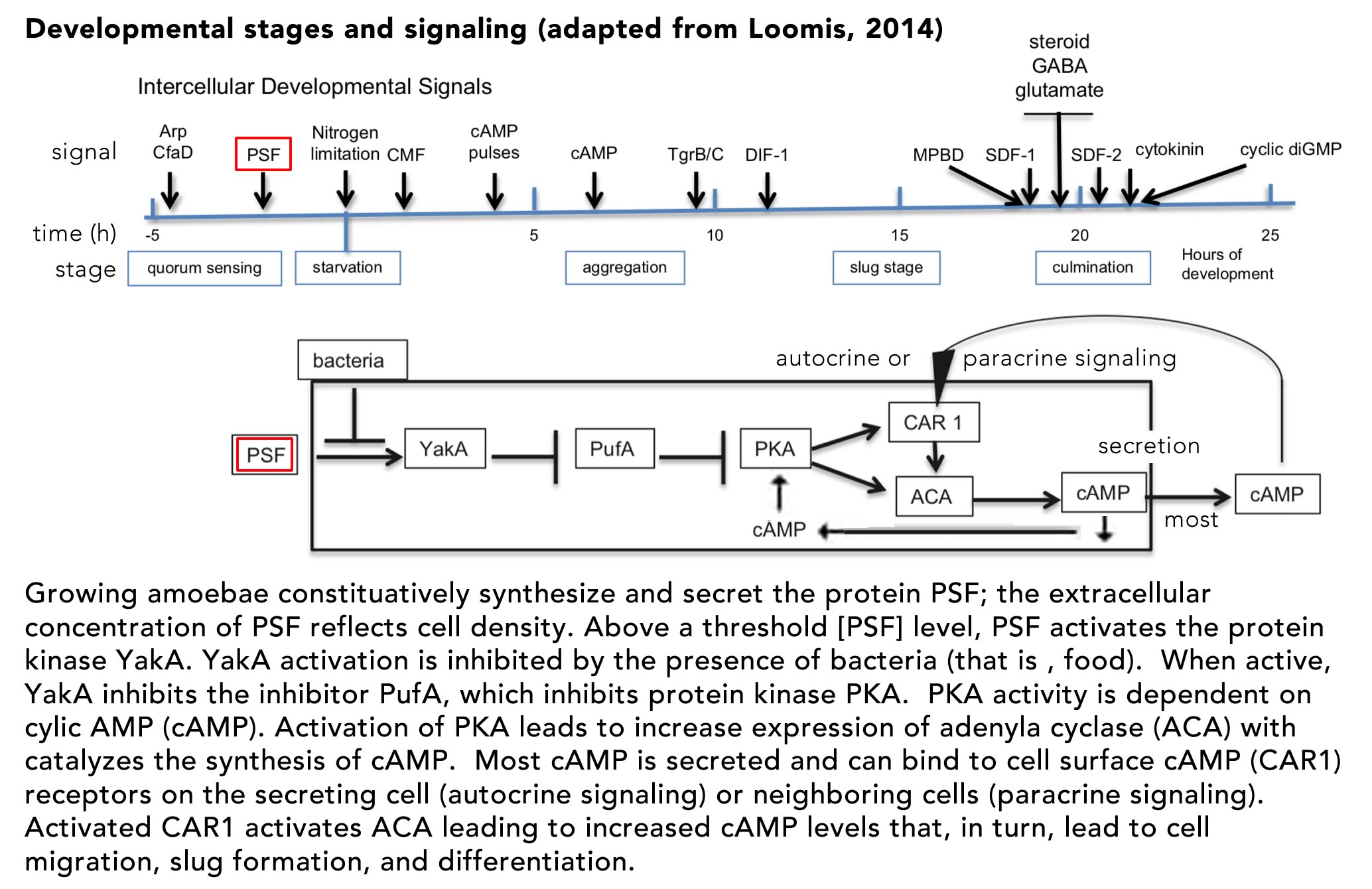
The process of cellular differentiation in D. discoideum has been worked out in molecular detail and involves two distinct signaling systems: the secreted pre-starvation factor (PSF) protein and cyclic AMP (cAMP).PSF is a quorum signaling protein that also serves to activate the cell aggregation and differentiation program (FIG. ↓)
If bacteria, that is food, are present, the activity of PSF is inhibited andcells remain in their single cell state. The key regulator of downstream aggregation and differentiation is the cAMP-dependent protein kinase PKA.In the unicellular state, PKA activity is inhibited by PufA.As PSF increases, while food levels decrease, YakA activity increases, inactivating PufA, leading to increased PKA activity.Active PKA induces the synthesis of two downstream proteins, adenylate cyclase (ACA) and the cAMP receptor (CAR1). ACA catalyzes cAMP synthesis, much of which is secreted from the cell as a signaling molecule. The membrane-bound CAR1 protein acts as a receptor for autocrine (on the cAMP secreting cell) and paracrine (on neighboring cells) signaling.The binding of cAMP to CAR1 leads to further activation of PKA, increasing cAMP synthesis and secretion – a positive feed-back loop. As cAMP levels increase, downstream genes are activated (and inhibited) leading cells to migrate toward one another, their adhesion to form a slug.Once the slug forms and migrates to an appropriate site, the process of differentiation (and death) leading to stalk and spore formation begins. The fates of the aggregated cells is determined stochastically, but social cheaters can arise. Mutations can lead to individuals that avoid becoming stalk cells.In the long run, if all individuals were to become cheaters, it would be impossible to form a stalk, so the purpose of social cooperation would be impossible to achieve.In the face of environmental variation, populations invaded by cheaters are more likely to become extinct.For our purposes the various defenses against cheaters are best left to other courses (see here if interested Strassmann et al., 2000).
Clonal (permanent) multicellularity:The type of multicellularity that most developmental biology courses focus on is what is termed clonal multicellularity – the organism is a clone of an original cell, the zygote, a diploid cell produced by the fusion of sperm and egg, haploid cells formed through the process of meiosis (2).It is during meiosis that most basic genetic processes occur, that is the recombination between maternal and paternal chromosomes leading to the shuffling of alleles along a chromosome, and the independent segregation of chromosomes to form haploid gametes, gametes that are genetically distinct from those present in either parent. Once the zygote forms, subsequent cell divisions involve mitosis, with only a subset of differentiated cells, the cells of the germ line, capable of entering meiosis.
Non-germ line, that is somatic cells, grow and divide. They interact with one another directly and through various signaling processes to produce cells with distinct patterns of gene expression, and so differentiated behaviors.A key difference from a unicellular organism, is that the cells will (largely) stay attached to one another, or to extracellular matrix materials secreted by themselves and their neighbors.The result is ensembles of cells displaying different specializations and behaviors.As such cellular colonies get larger, they face a number of physical constraints – for example, cells are open non-equilibrium systems, to maintain themselves and to grow and reproduce, they need to import matter and energy from the external world. Cells also produce a range of, often toxic, waste products that need to be removed.As the cluster of zygote-derived cells grows larger, and includes more and more cells, some cells will become internal and so cut off from necessary resources. While diffusive processes are often adequate when a cell is bathed in an aqueous solution, they are inadequate for a cell in the interior of a large cell aggregate (3).The limits of diffusive processes necessitate other strategies for resource delivery and waste removal; this includes the formation of tubular vascular systems (such as capillaries, arteries, veins) and contractile systems (hearts and such) to pump fluids through these vessels, as well as cells specialized to process and transport a range of nutrients (such as blood cells).As organisms get larger, their movements require contractile machines (muscle, cartilage, tendons, bones, etc) driving tails, fins, legs, wings, etc. The coordination of such motile systems involves neurons, ganglia, and brains. There is also a need to establish barriers between the insides of an organism and the outside world (skin, pulmonary, and gastrointestinal linings) and the need to protect the interior environment from invading pathogens (the immune system).The process of developing these various systems depends upon controlling patterns of cell growth, division, and specialization (consider the formation of an arm), as well as the controlled elimination of cells (apoptosis), important in morphogenesis (forming fingers from paddle-shaped appendages), the maturation of the immune system (eliminating cells that react against self), and the wiring up, and adaptation of the nervous system. Such changes are analogous to those involved in aggregative multicellularity.
Origins of multicellularity:While aggregative multicellularity involves an extension of quorum sensing and social cooperation between genetically distinct, but related individuals, we can wonder whether similar drivers are responsible for clonal multicellularity.There are a number of imaginable adaptive (evolutionary) drivers but two spring to mind: a way to avoid predators by getting bigger than the predators and as a way to produce varied structures needed to exploit various ecological niches and life styles. An example of the first type of driver of multicellularity is offered by the studies of Boraas et al(1998). They cultured the unicellular green alga Chlorella vulgaris, together with a unicellular predator, the phagotrophic flagellated protist Ochromonas vallescia. After less than 100 generations (cell divisions), they observed the appearance of multicellular, and presumable inedible (or at least less easily edible), forms. Once selected, this trait appears to be stable, such that “colonies retained the eight-celled form indefinitely in continuous culture”.To my knowledge, the genetic basis for this multicellularity remains to be determined.
Cell Differentiation:One feature of simple colonial organisms is that when dissociated into individual cells, each cell is capable of regenerating a new organism. The presence of multiple (closely related) cells in a single colony opens up the possibility of social interactions; this is distinct from the case in aggregative multicellularity, where social cooperation came first. Social cooperation within a clonal metazoan means that most cells “give up” their ability to reproduce a new organism (a process involving meiosis). Such irreversible social interactions mark the transition from a colonial organism to a true multicellular organism. As social integration increases, cells can differentiate so as to perform increasingly specialized functions, functions incompatible with cell division. Think for a moment about a human neuron or skeletal muscle cell – in both cases, cell division is no longer possible (apparently). Nevertheless, the normal functioning of such cells enhances the reproductive success of the organism as a whole – a classic example of inclusive fitness (remember heterocysts?)Modern techniques of single cell sequencing and data analysis have now been employed to map this process of cellular differentiation in increasingly great detail, observations that will inform our later discussions (see Briggs et al., 2018 and future posts). In contrast, the unregulated growth of a cancer cell is an example of an asocial behavior, an asocial behavior that is ultimately futile, except in those rare cases (four known at this point) in which a cancer cell can move from one organism to another (Ujvari et al., 2016).
Unicellular affordances for multicellularity:When considering the design of a developmental biology course, we are faced with the diversity of living organisms – the basic observation that Darwin, Wallace, their progenitors and disciplinary descendants set out to solve. After all there are many millions of different types of organisms; among the multicellular eukaryotes, there are six major group : the ascomycetes and basidiomycetes fungi, the florideophyte red algae, laminarialean brown algae, embryophytic land plants and animals
(Knoll, 2011 ↑).Our focus will be on animals. “All members of Animalia are multicellular, and all are heterotrophs (i.e., they rely directly or indirectly on other organisms for their nourishment). Most ingest food and digest it in an internal cavity.” [Mayer link].From a macroscopic perspective, most animals have (or had at one time during their development) an anterior to posterior, that is head to tail, axis. Those that can crawl, swim, walk, or fly typically have a dorsal-ventral or back to belly axis, and some have a left-right axis as well.
But to be clear, a discussion of the various types of animals is well beyond the scope of any introductory course in developmental biology, in part because there are 35 (assuming no more are discovered) different “types” (phyla) of animals – nicely illustrated at this website [BBC: 35 types of animals, most of whom are really weird)].So again, our primary focus will be on one group, the vertebrates – humans are members of this group.We will also consider experimental insights derived from studies of various “model” systems, including organisms from another metazoan group, theecdysozoa (organisms that shed their outer layer as they grow bigger), a group that includes fruit flies and nematode worms.
My goal will be to ignore most of the specialized terminology found in the scholarly literature, which can rapidly turn a biology course into a vocabulary lesson and that add little to understanding of basic processes relevant to a general understanding of developmental processes (and relevant to human biology, medicine, and biotechnology). This approach is made possible by the discovery that the basic processes associated with animal (and metazoan) development are conserved. In this light, no observation has been more impactful than the discovery that the nature and organization of the genes involved in specifying the head to tail axes of the fruit fly and vertebrates (such as the mouse and human) is extremely similar in terms of genomic organization and function (Lappin et al., 2006 ↓), an observation that we will return to repeatedly.Such molecular similarities extend to cell-cell and cell-matrix adhesion systems, systems that release and respond to various signaling molecules, controlling cell behavior and gene expression, and reflects the evolutionary conservation and the common ancestry of all animals (Brunet and King, 2017; Knoll, 2011).
What can we know about the common ancestor of the animals?Early on in the history of comparative cellular anatomy, the striking structural similarities between the feeding system of choanoflagellate protozoans, a motile (microtubule-based) flagellum a surrounded by a “collar”of microfilament-based microvilli) and a structurally similar organelle in a range of multicellular organisms led to the suggestion that choanoflagellates and animals shared a common ancestor.The advent of genomic sequencing and analysis has only strengthened this hypothesis, namely that choanoflagellates and animals form a unified evolutionary clade, the ‘Choanozoa’(see tree↑ above)(Brunet and King, 2017).Moreover, “many genes required for animal multicellularity (e.g., tyrosine kinases, cadherins, integrins, and extracellular matrix domains) evolved before animal origins”.The implications is that the Choanozoan ancestor was predisposed to exploit some of the early opportunities offered by clonal multicellularity. These pre-existing affordances, together with newly arising genes and proteins (Long et al., 2013) were exploited in multiple lineages in the generation of multicellular organisms (see Knoll, 2011).
Basically to understand what happened next, some ~600 million years ago or so, we will approach the various processes involved in the shaping of animal development.Because all types of developmental processes, including the unicellular to colonial transition, involve changes in gene expression, we will begin with the factors involved in the regulation of gene expression.
Footnotes: 1). Please excuse the inclusive plural, but it seems appropriate in the context of what I hope will be a highly interactive course.
2). I will explicitly ignore variants as (largely) distractions, better suited for more highly specialized courses. 3). We will return to this problem when (late in the course, I think) we will discuss the properties of induced pluripotent stem cell (iPSC) derived organoids.
Literature cited: Bonner, J. T. (1998). The origins of multicellularity. Integrative Biology: Issues, News, and Reviews: Published in Association with The Society for Integrative and Comparative Biology 1, 27-36.
Boraas, M. E., Seale, D. B. and Boxhorn, J. E. (1998). Phagotrophy by a flagellate selects for colonial prey: a possible origin of multicellularity. Evolutionary Ecology 12, 153-164.
Briggs, J. A., Weinreb, C., Wagner, D. E., Megason, S., Peshkin, L., Kirschner, M. W. and Klein, A. M. (2018). The dynamics of gene expression in vertebrate embryogenesis at single-cell resolution. Science 360, eaar5780.
Brunet, T. and King, N. (2017). The origin of animal multicellularity and cell differentiation. Developmental cell 43, 124-140.
Gardner, T. S., Cantor, C. R. and Collins, J. J. (2000). Construction of a genetic toggle switch in Escherichia coli. Nature 403, 339-342.
Hillmann, F., Forbes, G., Novohradská, S., Ferling, I., Riege, K., Groth, M., Westermann, M., Marz, M., Spaller, T. and Winckler, T. (2018). Multiple roots of fruiting body formation in Amoebozoa. Genome biology and evolution 10, 591-606.
Knoll, A. H. (2011). The multiple origins of complex multicellularity. Annual Review of Earth and Planetary Sciences 39, 217-239.
Lappin, T. R., Grier, D. G., Thompson, A. and Halliday, H. L. (2006). HOX genes: seductive science, mysterious mechanisms. The Ulster medical journal 75, 23.
Long, M., VanKuren, N. W., Chen, S. and Vibranovski, M. D. (2013). New gene evolution: little did we know. Annual review of genetics 47, 307-333.
Loomis, W. F. (2014). Cell signaling during development of Dictyostelium. Developmental biology 391, 1-16.
Strassmann, J. E., Zhu, Y. and Queller, D. C. (2000). Altruism and social cheating in the social amoeba Dictyostelium discoideum. Nature 408, 965-967.
Ujvari, B., Gatenby, R. A. and Thomas, F. (2016). Transmissible cancers, are they more common than thought? Evolutionary applications 9, 633-634.
On teaching developmental biology and trying to decide where to start: differentiation
Having considered the content of courses in chemistry [1] andbiology [2, 3], and preparing to teach developmental biology for the first time, I find myself reflecting on how such courses might be better organized.In my department, developmental biology (DEVO) has returned after a hiatus as the final capstone course in our required course sequence, and so offers an opportunity within which to examine what students have mastered as they head into their more specialized (personal) educational choices.Rather than describe the design of the course that I will be teaching, since at this point I am not completely sure what will emerge, what I intend to do (in a series of posts) is to describe, topic by topic, the progression of key concepts, the observations upon which they are based, and the logic behind their inclusion.
Modern developmental biology emerged during the mid-1800s from comparative embryology [4] and was shaped by the new cell theory (the continuity of life and the fact that all organisms are composed of cells and their products) and the ability of cells to differentiate, that is, to adopt different structures and behaviors [5].Evolutionary theory was also key.The role of genetic variation based on mutations and selection, in the generation of divergent species from common ancestors, explained why a single, inter-connected Linnaean (hierarchical) classification system (the phylogenic tree of life →) of organisms was possible and suggested that developmental mechanisms were related to similar processes found in their various ancestors.
So then, what exactly are the primary concepts behind developmental biology and how do they emerge from evolutionary, cell, and molecular biology?The concept of “development” applies to any process characterized by directional changes over time.The simplest such process would involve the progress from the end of one cell division event to the beginning of the next; cell division events provide a convenient benchmark.In asexual species, the process is clonal, a single parent gives rise to a genetically identical (except for the occurrence of new mutations) offspring. Often there is little distinction between parent and offspring.In sexual species, a dramatic and unambiguous benchmark involves the generation of a new and genetically distinct organism.This “birth” event is marked by the fusion of two gametes (fertilization) to form a new diploid organism.Typically gametes are produced by a complex cellular differentiation process (gametogenesis), ending with meiosis and the formation of haploid cells.In multicellular organisms, it is often the case that a specific lineage of cells (which reproduce asexually), known as the germ line, produce the gametes.The rest of the organism, the cells that do not produce gametes, is known as the soma, composed of somatic cells. Cellular continuity remains, however, since gametes are living (albeit haploid) cells.
It is common for the gametes that fuse to be of two different types, termed oocyte and sperm.The larger, and generally immotile gamete type is called an oocyte and an individual that produces oocytes is termed female. The smaller, and generally motile gamete type is called a sperm; individuals that produces sperm are termed male. Where a single organism can produce both oocytes and sperm, either at the same time or sequentially, they are referred to as hermaphrodites (named after Greek Gods, the male Hermes and the female Aphrodite). Oocytes and sperm are specialized cells; their formation involves the differential expression of genes and the specific molecular mechanisms that generate the features characteristic of the two cell types.The fusion of gametes, fertilization,leads to a zygote, a diploid cell that (usually) develops into a new, sexually mature organism.
An important feature of the process of fertilization is that it requires a level of social interaction, the two fusing cells (gametes) must recognize and fuse with one another.The organisms that produce these gametes must cooperate; they need to produce gametes at the appropriate time and deliver them in such a way that they can find and recognize each other and avoid “inappropriate” interactions”. The specificity of such interactions underlie the reproductive isolation that distinguishes one species from another.The development of reproductive isolation emerges as an ancestral population of organisms diverges to form one or more new species.As we will see, social interactions, and subsequent evolutionary effects, are common in the biological world.
The cellular and molecular aspects of development involve the processes by which cells grow, replicate their genetic material (DNA replication), divide to form distinct parent-offspring or similar sibling cells, and may alter their morphology (shape), internal organization, motility, and other behaviors, such as the synthesis and secretion of various molecules, and how these cells respond to molecules released by other cells.Developmental processes involve the expression and the control of all of these processes.
Essentially all changes in cellular behavior are associated with changes in the activities of biological molecules and the expression of genes, initiated in response to various external signaling events – fertilization itself is such a signal.These signals set off a cascade of regulatory interactions, often leading to multiple “cell types”, specialized for specific functions (such as muscle contraction, neural and/or hormonal signaling, nutrient transport, processing, and synthesis, etc.).For specific parts of the organism, external or internal signals can result in a short term “adaptive” response (such as sweating or panting in response to increased internal body temperature), after which the system returns to its original state, or in the case of developing systems, to new states, characterized by stable changes in gene expression, cellular morphology, and behavior.
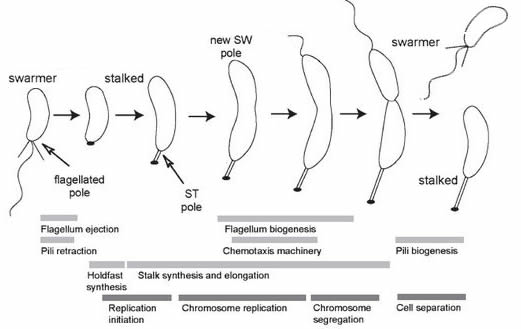
Development in bacteria (and other unicellular organisms):In most unicellular organisms, the cell division process is reasonably uneventful, the cells produced are similar to the original cell – but not always.A well studied example is the bacterium Caulobacter crescentus (and related species) [link][link].In cases such as this, the process of growthleads to phenotypically different daughters.While it makes no sense to talk about a beginning (given the continuity of life after the appearance of the last universal common ancestor or LUCA), we can start with a “swarmer” cell, characterized by the presence of a motile flagellum (a molecular machine driven by coupled chemical reactions – see past blogpost] that drives motility [figure modified from 6 ↓].
A swarmer will eventually settle down, loose the flagellum, and replace it with a specialized structure (a holdfast) designed to anchor the cell to a solid substrate.As the organism grows, the holdfast develops a stalk that lifts the cell away from the substrate.As growth continues, the end of the cell opposite the holdfast begins to differentiate (becomes different) from the holdfast end of the cell – it begins the process leading to the assembly of a new flagellar apparatus.When reproduction (cell growth, DNA replication, and cell division) occurs, a swarmer cell is released and can swim away and colonize another area, or settle nearby.The holdfast-anchored cell continues to grow, producing new swarmers. This process is based on the inherent asymmetry of the system – the holdfast end of the cell is molecularly distinct from the flagellar end [see 7].
The process of swarmer cell formation in Caulobacter is an example of what we will term deterministic phenotypic switching.Cells can also exploit molecular level noise (stochastic processes) that influence gene expression to generate phenotypic heterogeneity, different behaviors expressed by genetically identical cells within the same environment [see 8, 9].Molecular noise arises from the random nature of molecular movements and the rather small (compared to macroscopic systems) numbers of most molecules within a cell.Most cells contain one or two copies of any particular gene, and a similarly small number of molecular sequences involved in their regulation [10].Which molecules are bound to which regulatory sequence, and for how long, is governed by inter-molecular surface interactions and thermally driven collisions, and is inherently noisy.There are strategies that can suppress but not eliminate such noise [see 11].As dramatically illustrated by Elowitzand colleagues [8](↑), molecular level noise can produce cells with different phenotypes.Similar processes are active in eukaryotes (including humans), and can lead to the expression of one of the two copies of a gene (mono-allelic expression) present in a diploid organism.This can lead to effects such as haploinsufficiency and selective (evolutionary) lineage effects if the two alleles are not identical [12, 13]. Such phenotypic heterogeneity among what are often genetically identical cells is a topic that is rarely discussed (as far as I can discern) in introductory cell, molecular, or developmental biology courses [past blogpost].
The ability to switch phenotypes can be a valuable trait if an organism’s environment is subject to significant changes.As an example, when the environment gets hostile, some bacterial cells transition from a rapidly dividing to a slow or non-dividing state.Such “spores” can differentiate so as to render them highly resistant to dehydration and other stresses.If changes in environment are very rapid, a population can protect itself by continually having some cells (stochastically) differentiating into spores, while others continue to divide rapidly. Only a few individuals (spores) need to survive a catastrophic environmental change to quickly re-establish the population.
Dying for others – social interactions between “unicellular” organisms:Many students might not predict that one bacterial cell would “sacrifice” itself for the well being of others, but in fact there are a number of examples of this type of self-sacrificing behavior, known as programmed cell death, which is often a stochastic process.An interesting example is provided by cellular specialization for photosynthesis or nitrogen fixation in cyanobacteria [see 9].These two functions require mutually exclusive cellular environments to occur, in particular the molecular oxygen (O2) released by photosynthesis inhibits the process of nitrogen fixation.Nevertheless, both are required for optimal growth.The solution?some cells differentiate into what are known as heterocysts, cells committed to nitrogen fixation (← a heterocyst in Anabaena spiroides, adapted from link), while most ”vegetative” cells continue with photosynthesis.Heterocysts cannot divide, and eventually die – they sacrifice themselves for the benefit of their neighbors, the vegetative cells, cells that can reproduce.
The process by which the death of an individual can contribute resources that can be used to insure or enhance the survival and reproduction of surrounding individuals is an inherently social process, and is subject of social evolutionary mechanisms [14, 15][past blogpost]. Social behaviors can be selected for because the organism’s neighbors, the beneficiaries of their self-sacrifice are likely to be closely (clonally) related to themselves. One result of the social behavior is, at the population level, an increase in one aspect of evolutionary fitness,termed “inclusive fitness.”
Such social behaviors can enable a subset of the population to survive various forms of environmental stress (see spore formation above).An obvious environmental stress involves the impact of viral infection.Recall that viruses are completely dependent upon the metabolic machinery of the infected cell to replicate. While there are a number of viral strategies, a common one is bacterial lysis – the virus replicates explosively, kills the infected cells, leading to the release of virus into the environment to infect others.But, what if the infected cell kills itself BEFORE the virus replicates – the dying (self-sacrificing, altruistic) cell “kills” the virus (although viruses are not really alive) and stops the spread of the infection.Typically such genetically programmed cell death responses are based on a simple two-part system, involving a long lived toxin and a short-lived anti-toxin.When the cell is stressed, for example early during viral infection, the level of the anti-toxin can fall, leading to the activation ofthe toxin.
Other types of social behavior and community coordination (quorum effects): Some types of behaviors only make sense when the density of organisms rises above a certain critical level.For example,it would make no sense for an Anabaena cellto differentiate into a heterocyst (see above) if there are no vegetative cells nearby.Similarly, there are processes in which a behavior of a single bacterial cell, such as the synthesis and secretion of a specific enzyme, a specific import or export machine, or the construction of a complex, such as a DNA uptake machine, makes no sense in isolation – the secreted molecule will just diffuse away, and so be ineffective, the molecule to be imported (e.g. lactose) or exported (an antibiotic) may not be present, or there may be no free DNA to import.However, as the concentration (organisms per volume) of bacteria increases, these behaviors can begin to make biological sense – there is DNA to eat or incorporate and the concentration of secreted enzyme can be high enough to degrade the target molecules (so they are inactivated or can be imported as food).
So how does a bacterium determine whether it has neighbors or whether it wants to join a community of similar organisms?After all, it does not have eyes to see. The process used is known as quorum sensing.Each individual synthesizes and secretes a signaling molecule and a receptor protein whose activity is regulated by the binding of the signaling molecule. Species specificity in signaling molecules and receptors insures that organisms of the same kind are talking to one another and not to other, distinct types of organisms that may be in the environment. At low signaling molecule concentrations, such as those produced by a single bacterium in isolation, the receptor is not activated and the cell’s behavior remains unchanged.However, as the concentration of bacteria increases, the concentration of the signal increases, leading to receptor activation. Activation of the receptor can have a number of effects, including increased synthesis of the signal and other changes, such as movement in response to signals through regulation of flagellar and other motility systems, such a system can lead to the directed migration (aggregation) of cells [see 16].
In addition to driving the synthesis of a common good (such as a useful extracellular molecule), social interactions can control processes such as programmed cell death. When the concentration of related neighbors is high, the programmed death of an individual can be beneficial, it can lead to release of nutrients (common goods, including DNA molecules) that can be used by neighbors (relatives)[17, 18] – an increase in the probability of cell death in response to a quorum can increased in a way that increases inclusive fitness.On the other hand,if there are few related individuals in the neighborhood, programmed cell death “wastes” these resources, and so is likely to be suppressed (you might be able to generate a plausible mechanism that could control the probability of programmed cell death).
As we mentioned previously with respect to spore formation, the generation of a certain percentage of “persisters” – individuals that withdraw from active growth and cell division, can enable a population to survive stressful situations, such as the presence of an antibiotic. On the other hand, generating too many persisters may place the population at a reproductive disadvantage. Once the antibiotic is gone, the persisters can return into active division. The ability of bacteria to generate persisters is a serious problem in treating people with infections, particularly those who stop taking their antibiotics too early [19].
Of course, as in any social system, the presumption of cooperation (expending energy to synthesize the signal, sacrificing oneself for others) can open the system to cheaters [blogpost].All such “altruistic” behaviors are vulnerable to cheaters.*For example, a cheater that avoids programmed cell death (for example due to an inactivating mutation that effects the toxin molecule involved) will come to take over the population.The downside, for the population, is that if cheaters take over, the population is less likely to survive the environmental events that the social behavior was evolve to address.In response to the realities of cheating, social organisms adopt various social-validation and policing systems [see 20 as an example]; we see this pattern of social cooperation, cheating, and social defense mechanism throughout the biological world.
* Such as people who fail to pay their taxes or disclose their tax returns.
literature cited:
1. Cooper, M.M. and M.W. Klymkowsky, Chemistry, life, the universe, and everything: a new approach to general chemistry, and a model for curriculum reform. J. Chem. Educ. 2013. 90: 1116-1122 & Cooper, M. M., R. Stowe, O. Crandell and M. W. Klymkowsky. Organic Chemistry, Life, the Universe and Everything (OCLUE): A Transformed Organic Chemistry Curriculum. J. Chem. Educ. 2019. 96: 1858-1872.
2. Klymkowsky, M.W., Teaching without a textbook: strategies to focus learning on fundamental concepts and scientific process. CBE Life Sci Educ, 2007. 6: 190-3.
3. Klymkowsky, M.W., J.D. Rentsch, E. Begovic, and M.M. Cooper, The design and transformation of Biofundamentals: a non-survey introductory evolutionary and molecular biology course. LSE Cell Biol Edu, 2016. pii: ar70.
4. Arthur, W., The emerging conceptual framework of evolutionary developmental biology. Nature, 2002. 415: 757.
5. Wilson, E.B., The cell in development and heredity. 1940.
6. Jacobs‐Wagner, C., Regulatory proteins with a sense of direction: cell cycle signalling network in Caulobacter. Molecular microbiology, 2004. 51:7-13.
7. Hughes, V., C. Jiang, and Y. Brun, Caulobacter crescentus. Current biology: CB, 2012. 22:R507.
8. Elowitz, M.B., A.J. Levine, E.D. Siggia, and P.S. Swain, Stochastic gene expression in a single cell. Science, 2002. 297:1183-6.
9. Balázsi, G., A. van Oudenaarden, and J.J. Collins, Cellular decision making and biological noise: from microbes to mammals. Cell, 2011. 144: 910-925.
10. Fedoroff, N. and W. Fontana, Small numbers of big molecules. Science, 2002. 297:1129-1131.
11. Lestas, I., G. Vinnicombe, and J. Paulsson, Fundamental limits on the suppression of molecular fluctuations. Nature, 2010. 467:174-178.
12. Zakharova, I.S., A.I. Shevchenko, and S.M. Zakian, Monoallelic gene expression in mammals. Chromosoma, 2009. 118:279-290.
13. Deng, Q., D. Ramsköld, B. Reinius, and R. Sandberg, Single-cell RNA-seq reveals dynamic, random monoallelic gene expression in mammalian cells. Science, 2014. 343: 193-196.
14. West, S.A., A.S. Griffin, A. Gardner, and S.P. Diggle, Social evolution theory for microorganisms. Nature reviews microbiology, 2006. 4:597.
15. Bourke, A.F.G., Principles of Social Evolution. Oxford series in ecology and evolution. 2011, Oxford: Oxford University Press.
16. Park, S., P.M. Wolanin, E.A. Yuzbashyan, P. Silberzan, J.B. Stock, and R.H. Austin, Motion to form a quorum. Science, 2003. 301:188-188.
17. West, S.A., S.P. Diggle, A. Buckling, A. Gardner, and A.S. Griffin, The social lives of microbes. Annual Review of Ecology, Evolution, and Systematics, 2007: 53-77.
18. Durand, P.M. and G. Ramsey, The Nature of Programmed Cell Death. Biological Theory, 2018: 1-12.
19. Fisher, R.A., B. Gollan, and S. Helaine, Persistent bacterial infections and persister cells. Nature Reviews Microbiology, 2017. 15:453.
20. Queller, D.C., E. Ponte, S. Bozzaro, and J.E. Strassmann, Single-gene greenbeard effects in the social amoeba Dictyostelium discoideum. Science, 2003. 299: 105-106.
Links between genetics and race crop up periodically in the popular press (link; link), but the real, substantive question, and the topic of a number of recent essays (see Saletan. 2018a. Stop Talking About Race and IQ) is whether the idea of “race” as commonly understood, and used by governments to categorize people (link), makes scientific sense.More to the point, do biology educators have an unmet responsibility to modify and extend their materials and pedagogical approaches to address the non-scientific, often racist, implications of racial characterizations.Such questions are complicated by a second factor, independent of whether the term race has any useful scientific purpose, namely to help students understand the biological (evolutionary) origins of racism itself, together with the stressors that lead to its periodic re-emergence as a socio-political factor. In times of social stress, reactions to strangers (others) identified by variations in skin color or overt religious or cultural signs (dress), can provoke hostility against those perceived to be members of a different social group.As far as I can tell, few in the biology education community, which includes those involved in generating textbooks, organizing courses and curricula, or the design, delivery, and funding of various public science programs, including PBS’s NOVA, the science education efforts of HHMI and other private foundations, and programs such as Science Friday on public radio, directly address the roots of racism, roots associated with biological processes such as the origins and maintenance of multicellularity and other forms of social organization among organisms, involved in coordinating their activities and establishing defenses against social cheaters and processes such as cancer, in an organismic context (1).These established defense mechanisms can, if not recognized and understood, morph into reflexive and unjustified intolerance, hostility toward, and persecution of various “distinguishable others.”I will consider both questions, albeit briefly, here.
Two factors have influenced my thinking about these questions.The first involves the design of the biofundamentals text/course and its extension to include topics in genetics (2).This involved thinking about what is commonly taught in genetics, what is critical for students to know going forward (and by implication what is not), and where materials on genetic processes best fit into a molecular biology curriculum (3).While engaged in such navel gazing there came an email from Malcolm Campbell describing student responses to the introduction of a chapter section on race and racism in his textbook Integrating Concepts in Biology.The various ideas of race, the origins of racism, and the periodic appearance of anti-immigrant, anti-religious and racist groups raise important questions – how best to clarify what is an undeniable observation, that different, isolated, sub-populations of a species can be distinguished from one another (see quote from Ernst Mayr’s 1994 “Typological versus Population thinking” →), from the deeper biological reality, that at the level of the individual these differences are meaningless. In what I think is an interesting way, the idea that people can be meaningfully categorized as different types of various platonic ideals (for example, as members of one race or the other) based on anatomical / linguistic differences between once distinct sub-populations of humans is similar to the dichotomy between common wisdom (e.g. that has influenced people’s working understanding of the motion of objects) and the counter-intuitive nature of empirically established scientific ideas (e.g. Newton’s laws and the implications of Einstein’s theory of general relativity).What appears on the surface to be true but in fact is not.In this specific case, there is a pressure toward what Mayr terms “typological” thinking, in which we class people into idealized (platonic) types or races (↓).
As pointed out most dramatically, and repeatedly, by Mayr (1985; 1994; 2000), and supported by the underlying commonality of molecular biological mechanisms and the continuity of life, stretching back to the last universal common ancestor, there are only individuals who are members of various populations that have experienced various degrees of separation from one another.In many cases, these populations have diverged and, through geographic, behavioral, and structure adaptations driven by natural, social, and sexual selection together with the effects of various events, some non-adaptive, such as bottlenecks, founder effects, and genetic drift, may eventually become reproductively isolated from one another, forming new species.An understanding of evolutionary principles and molecular mechanisms transforms biology from a study of non-existent types to a study of populations with their origins in common, sharing a single root – the last universal common ancestor (LUCA). Over the last ~200,000 years the movement of humans first within Africa and then across the planet has been impressive (→). These movements have been accompanied by the fragmentation of human populations. Campbell and Tishkoff (2008) identified 13 distinct ancestral African populations while Busby et al (2016) recognized 48 sub-saharan population groups. The fragmentation of the human population is being reversed (or rather rendered increasingly less informative) by the effects of migration and extensive intermingling (↓).
Ideas, such as race (and in a sense species), try to make sense of the diversity of the many different types of organisms we observe. They are based on a form of essentialist or typological thinking – thinking that different species and populations are completely different “kinds” of objects, rather than individuals in a population connected historically to all other living things. Race is a more pernicious version of this illusion, a pseudo-scientific, political and ideological idea that postulates that humans comein distinct, non-overlapping types (quote ↓ again, from Mayr).Such a weird idea underlies various illogical and often contradictory legal “rules” by which a person’s “race” is determined.
Given the reality of the individual and the unreality of race, racial profiling (see Satel,
2002) can lead to serious medical mistakes, as made clear in the essays by Acquaviva & Mintz (2010) “Are We Teaching Racial Profiling?”, Yudell et al(2016) “Taking Race out of Human Genetics”, and Donovan (2014) “The impact of the hidden curriculum”.
The idea of race as a type fails to recognize the dynamics of the genome over time.If possible (sadly not) a comparative analysis of the genome of a “living fossil”, such as modern day coelacanths and their ancestors (living more than 80 million years ago) would likely reveal dramatic changes in genomic DNA sequence.In this light the fact that between 100 to 200 new mutations are introduced into the human genome per generation (see Dolgin 2009 Human mutation rate revealed) seems like a useful number to be widely appreciated by students, not to mention the general public. Similarly, the genomic/genetic differences between humans, our primate relatives, and other mammals and the mechanisms behind them (Levchenko et al., 2017)(blog link) would seem worth considering and explicitly incorporating into curricula on genetics and human evolution.
While race may be meaningless, racism is not.How to understand racism?Is it some kind of political artifact, or does it arise from biological factors.Here, I believe, we find a important omission in many biology courses, textbooks, and curricula – namely an introduction and meaningful discussion of social evolutionary mechanisms. Many is the molecular/cell biology curriculum that completely ignores such evolutionary processes. Yet, the organisms that are the primary focus of biological research (and who pay for such research, e.g. humans) are social organisms at two levels.In multicellular organisms somatic cells, which specialize to form muscular, neural, circulatory and immune systems, bone and connective tissues, sacrifice their own inter-generational reproductive future to assist their germ line (sperm and/or eggs) relatives, the cells that give rise to the next generation of organisms, a form of inclusive fitness (Dugatkin, 2007).Moreover, humans are social organisms, often sacrificing themselves, sharing their resources, and showing kindness to other members of their group. This social cooperation is threatened by cheaters of various types (POST LINK).Unless these social cheaters are suppressed, by a range of mechanisms, and through processes of kin/group selection, multicellular organisms die and socially dysfunctional social populations are likely to die out.Without the willingness to cooperate, and when necessary, self-sacrifice, social organization is impossible – no bee hives, no civilizations.Imagine a human population composed solely of people who behave in a completely selfish manner, not honoring their promises or social obligations.
A key to social interactions involves recognizing those who are, and who are not part of your social group.A range of traits can serve as markers for social inclusion.A plausible hypothesis is that the explicit importance of group membership and defined social interactions becomes more critical when a society, or a part of society, is under stress.Within the context of social stratification, those in the less privileged groups may feel that the social contract has been broken or made a mockery of.The feeling (apparent reality) that members of “elite” or excessively privileged sub-groups are not willing to make sacrifices for others serves as evidence that social bonds are being broken (4). Times of economic and social disruption (migrations and conquests) can lead to increased explicit recognition of both group and non-group identification.The idea that outsiders (non-group members) threaten the group can feed racism, a justification for why non-group members should be treated differently from group members.From this position it is a small (conceptual) jump to the conclusion that non-group members are somehow less worthy, less smart, less trustworthy, less human – different in type from members of the group – many of these same points are made in an op-ed piece by Judis. 2018. What the Left Misses About Nationalism.
That economic or climatic stresses can foster the growth of racist ideas is no new idea; consider the unequal effects of various disruptions likely to be associated with the spread ofautomation (quote from George Will↓) and the impact of climate change on migrations of groups within and between countries (see Saletan 2018b: Why Immigration Opponents Should Worry About Climate Change) are likely to spur various forms of social unrest, whether revolution or racism, or both – responses that could be difficult to avoid or control.
So back to the question of biology education – in this context understanding the ingrained responses of social creatures associated with social cohesion and integrity need to be explicitly presented. Similarly, variants of such mechanisms occur within multicellular organisms and how they work is critical to understanding how diseases such as cancer, one of the clearest forms of a cheater phenotype, are suppressed.Social evolutionary mechanisms provide the basis for understanding a range of phenomena, and the ingrained effects of social selection may be seen as one of the roots of racism, or at the very least a contributing factor worth acknowledging explicitly.
Thanks to Melanie Cooper and Paul Strode for comments. Minor edits 4 May 2019.
Footnotes:
It is an interesting possibility whether the 1%, or rather the super 0.1% represent their own unique form of social parasite, leading periodically to various revolutions – although sadly, new social parasites appear to re-emerge quite quickly.
At this point it is worth noting that biofundamentals itself includes sections on social evolution, kin/group and sexual selection (see Klymkowsky et al., 2016; LibreText link).
One might be forgiven for thinking that rich and privileged folk who escape paying what is seen as their fair share of taxes, might be cast as social cheaters (parasites) who, rather than encouraging racism might lead to revolutionary thoughts and actions.
Literature cited:
Acquaviva & Mintz. (2010). Perspective: Are we teaching racial profiling? The dangers of subjective determinations of race and ethnicity in case presentations. Academic Medicine85, 702-705.
Busby et al. (2016). Admixture into and within sub-Saharan Africa. Elife5, e15266.
Campbell & Tishkoff. (2008). African genetic diversity: implications for human demographic history, modern human origins, and complex disease mapping. Annu. Rev. Genomics Hum. Genet.9, 403-433.
Donovan, B.M. (2014). Playing with fire? The impact of the hidden curriculum in school genetics on essentialist conceptions of race. Journal of Research in Science Teaching51: 462-496.
Dugatkin, L. A. (2007). Inclusive fitness theory from Darwin to Hamilton. Genetics176, 1375-1380.
Levchenko et al., (2017). Human accelerated regions and other human-specific sequence variations in the context of evolution and their relevance for brain development. Genome biology and evolution10, 166-188.
Mayr, E. (1985). The Growth of Biological Thought: Diversity, Evolution, and Inheritance. Cambridge, MA: Belknap Press of Harvard University Press.
Mayr, E. (1994). Typological versus population thinking. Conceptual issues in evolutionary biology, 157-160.
—- (2000). Darwin’s influence on modern thought. Scientific American283, 78-83.
Satel, S. (2002). I am a racially profiling doctor. New York Times5, 56-58.
Yudell et al., (2016). Taking race out of human genetics. Science351, 564-565.










 second factor, independent of whether the term race has any useful scientific purpose, namely to help students understand the biological (evolutionary) origins of racism itself, together with the stressors that lead to its periodic re-emergence as a socio-political factor. In times of social stress, reactions to strangers (others) identified by variations in skin color or overt religious or cultural signs (dress), can provoke hostility against those perceived to be members of a different social group.
second factor, independent of whether the term race has any useful scientific purpose, namely to help students understand the biological (evolutionary) origins of racism itself, together with the stressors that lead to its periodic re-emergence as a socio-political factor. In times of social stress, reactions to strangers (others) identified by variations in skin color or overt religious or cultural signs (dress), can provoke hostility against those perceived to be members of a different social group.





{kind=link}