
originally published as https://facultyopinions.com/article/739916951 – July 2021
There have been many calls for improved “scientific literacy”. Scientific literacy has been defined in a number of, often ambiguous, ways (see National Academies of Sciences and Medicine, 2016 {1}). According to Krajcik & Sutherland (2010) {2} it is “the understanding of science content and scientific practices and the ability to use that knowledge”, which implies “the ability to critique the quality of evidence or validity of conclusions about science in various media, including newspapers, magazines, television, and the Internet”. But what types of critiques are we talking about, and how often is this ability to critique, and the scientific knowledge it rests on, explicitly emphasized in the courses non-science (or science) students take? As an example, highlighted by Sabine Hossenfelder (2020) {3}, are students introduced to the higher order reasoning and understanding of the scientific enterprise needed to dismiss a belief in a flat (or a ~6000 year old) Earth?
While the sources of scientific illiteracy are often ascribed to social media, religious beliefs, or economically or politically motivated distortions, West and Bergstrom point out how scientists and the scientific establishment (public relations departments and the occasional science writer) also play a role. They identify the problems arising from the fact that the scientific enterprise (and the people who work within it) act within “an attention economy” and “compete for eyeballs just as journalists do.” The authors provide a review of all of the factors that contribute to misinformation within the scientific literature and its media ramifications, including the contribution of “predatory publishers” and call for “better ways of detecting untrustworthy publishers.” At the same time, there are ingrained features of the scientific enterprise that serve to distort the relevance of published studies, these include not explicitly identifying the organism in which the studies are carried out, and so obscuring the possibility that they might not be relevant to humans (see Kolata, 2013 {4}). There are also systemic biases within the research community. Consider the observation, characterized by Pandey et al. (2014) {5} that studies of “important” genes, expressed in the nervous system, are skewed: the “top 5% of genes absorb 70% of the relevant literature” while “approximately 20% of genes have essentially no neuroscience literature”. What appears to be the “major distinguishing characteristic between these sets of genes is date of discovery, early discovery being associated with greater research momentum—a genomic bandwagon effect”, a version of the “Matthew effect” described by Merton (1968) {6}. In the context of the scientific community, various forms of visibility (including pedigree and publicity) are in play in funding decisions and career advancement. Not pointed out explicitly by West and Bergstrom is the impact of disciplinary experts who pontificate outside of their areas of expertise and speculate beyond what can be observed or rejected experimentally, including speculations on the existence of non-observable multiverses, the ubiquity of consciousness (Tononi & Koch, 2015 {7}), and the rejection of experimental tests as a necessary criterion of scientific speculation (see Loeb, 2018 {8}) spring to mind.
Many educational institutions demand that non-science students take introductory courses in one or more sciences in the name of cultivating “scientific literacy”. This is a policy that seems to me to be tragically misguided, and perhaps based more on institutional economics than student learning outcomes. Instead, a course on “how science works and how it can be distorted” would be more likely to move students close to the ability to “critique the quality of evidence or validity of conclusions about science”. Such a course could well be based on an extended consideration of the West and Bergstrom article, together with their recently published trade book “Calling bullshit: the art of skepticism in a data-driven world” (Bergstrom and West, 2021 {9}), which outlines many of the ways that information can be distorted. Courses that take this approach to developing a skeptical (and realistic) approach to understanding how the sciences work are mentioned, although what measures of learning outcomes have been used to assess their efficacy are not described.
literature cited
- Science literacy: concepts, contexts, and consequencesCommittee on Science Literacy and Public Perception of Science, Board on Science Education, Division of Behavioral and Social Sciences and Education, National Academies of Sciences, Engineering, and Medicine.2016 10 14; PMID: 27854404
- Supporting students in developing literacy in science. Krajcik JS, Sutherland LM.Science. 2010 Apr 23; 328(5977):456-459PMID: 20413490
- Flat Earth “Science”: Wrong, but not Stupid. Hossenfelder S. BackRe(Action) blog, 2020, Aug 22 (accessed Jul 29, 2021)
- Mice fall short as test subjects for humans’ deadly ills. Kolata G. New York Times, 2013, Feb 11 (accessed Jul 29, 2021)
- Functionally enigmatic genes: a case study of the brain ignorome. Pandey AK, Lu L, Wang X, Homayouni R, Williams RW.PLoS ONE. 2014; 9(2):e88889PMID: 24523945
- The Matthew Effect in Science: The reward and communication systems of science are considered.Merton RK.Science. 1968 Jan 5; 159:56-63 PMID: 17737466
- Consciousness: here, there and everywhere? Tononi G, Koch C.Philos Trans R Soc Lond B Biol Sci. 2015 May 19; 370(1668)PMID: 25823865
- Theoretical Physics Is Pointless without Experimental Tests. Loeb A. Scientific American blog, 2018, Aug 10 [ Blog piece] (accessed Jul 29, 2021)
- Calling bullshit: the art of skepticism in a data-driven world.Bergstrom CT, West JD. Random House Trade Paperbacks, 2021ISBN: 978-0141987057

 second factor, independent of whether the term race has any useful scientific purpose, namely to help students understand the biological (evolutionary) origins of racism itself, together with the stressors that lead to its periodic re-emergence as a socio-political factor. In times of social stress, reactions to strangers (others) identified by variations in skin color or overt religious or cultural signs (dress), can provoke hostility against those perceived to be members of a different social group.
second factor, independent of whether the term race has any useful scientific purpose, namely to help students understand the biological (evolutionary) origins of racism itself, together with the stressors that lead to its periodic re-emergence as a socio-political factor. In times of social stress, reactions to strangers (others) identified by variations in skin color or overt religious or cultural signs (dress), can provoke hostility against those perceived to be members of a different social group.




 people as disposable machines, to be sacrificed for some ideological or religious faith (3).
people as disposable machines, to be sacrificed for some ideological or religious faith (3). This is not to imply that humans (and other animals) are totally in control of their thoughts and actions, completely “free” – obviously not.
This is not to imply that humans (and other animals) are totally in control of their thoughts and actions, completely “free” – obviously not.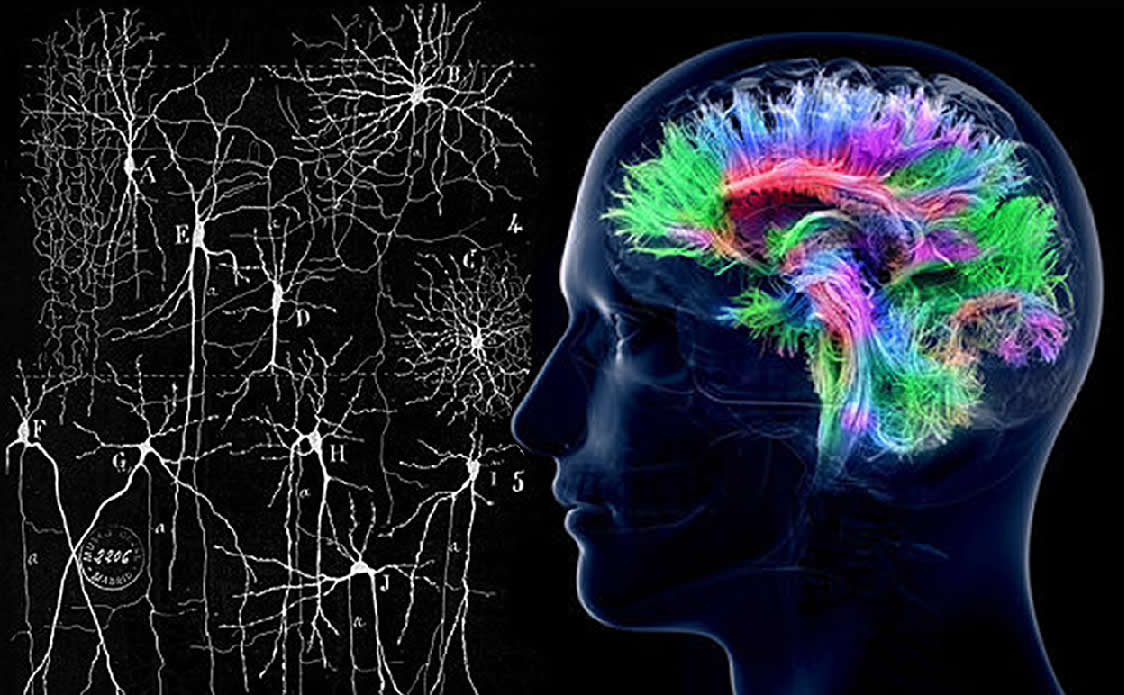 making ~7,000,000,000,000,000 synapses with one another, and interacting in various ways with ~100,000,000,000 glia that include non-neuronal astrocytes, oligodendrocytes, and immune system microglia (von Bartheld et al., 2016). These considerations ignore the recently discovered effects of the rest of the body (and its microbiome) on the brain (see Mayer et al., 2014; Smith, 2015).
making ~7,000,000,000,000,000 synapses with one another, and interacting in various ways with ~100,000,000,000 glia that include non-neuronal astrocytes, oligodendrocytes, and immune system microglia (von Bartheld et al., 2016). These considerations ignore the recently discovered effects of the rest of the body (and its microbiome) on the brain (see Mayer et al., 2014; Smith, 2015).


{kind=link}