
21st Century DEVO-2 In the first post in this series [link], I introduced the observation that single celled organisms can change their behaviors, often in response to social signals. They can respond to changing environments and can differentiate from one cellular state to the another. Differentiation involves changes in which sets of genes are expressed, which polypeptides and proteins are made [previous post], where the proteins end up within the cell, and which behaviors are displayed by the organism. Differentiation enables individuals to adapt to hostile conditions and to exploit various opportunities.
The ability of individuals to cooperate with one another, through processes such as quorum sensing, enables them to tune their responses so that they are appropriate and useful. Social interactions also makes it possible for them to produce behaviors that would be difficult or impossible for isolated individuals. Once individual organisms learn, evolutionarily, how to cooperate, new opportunities and challenges (cheaters) emerge. There are strategies that can enable an organism to adapt to a wider range of environments, or to become highly specialized to a specific environment, through the production of increasingly complex behaviors. As described previously, many of these cooperative strategies can be adopted by single celled organisms, but others require a level of multicellularity. Multicellularity can be transient – a pragmatic response to specific conditions, or it can be (if we ignore the short time that gametes exist as single cells) permanent, allowing the organism to develop the range of specialized cells types needed to build large, macroscopic organisms with complex and coordinated behaviors. In appears that various forms of multicellularity have arisen independently in a range of lineages (Bonner, 1998; Knoll, 2011). We can divide multicellularity into two distinct types, aggregative and clonal – which we will discuss in turn (1). Aggregative (transient) multicellularity: Once organisms had developed quorum sensing, they can monitor the density of related organisms in their environment and turn or (or off) specific genes (or sets of genes, necessary to produce a specific behavior. While there are many variants, one model for such a behavior is a genetic toggle switch, in which a particular gene (or genes) can be switched on or off in response to environmental signals acting as allosteric regulators of transcription factor proteins (see Gardner et al., 2000). Here is an example of an activity (↓) that we will consider in class to assess our understanding of the molecular processes involved.

One outcome of such a signaling system is to provoke the directional migration of amoeba and their aggregation to form the transient multicellular “slug”. Such behaviors has been observed in a range of normally unicellular organisms (see Hillmann et al., 2018)(↓). The classic example is the cellular slime mold Dictyostelium discoideum (Loomis, 2014). Under normal conditions, these

unicellular amoeboid eukaryotes migrate, eating bacteria and such. In this state, the range of an individual’s movement is restricted to short distances. However when conditions turn hostile, specifically a lack of necessary nitrogen compounds, there is a compelling reason to abandon one environment and migrate to another, more distant that a single-celled organism could reach. This is a behavior that depends upon the presence of a sufficient density (cells/unit volume) of cells that enables them to: 1) recognize one another’s presence (through quorum sensing), 2) find each other through directed (chemotactic) migration, and 3) form a multicellular slug that can go on to differentiate. Upon differentiation about 20% of the cells differentiate (and die), forming a stalk that lifts the other ~80% of the cells into the air. These non-stalk cells (the survivors) differentiate into spore (resistant to drying out) cells that are released into the air where they can be carried to new locations, establishing new populations.

The process of cellular differentiation in D. discoideum has been worked out in molecular detail and involves two distinct signaling systems: the secreted pre-starvation factor (PSF) protein and cyclic AMP (cAMP). PSF is a quorum signaling protein that also serves to activate the cell aggregation and differentiation program (FIG. ↓)
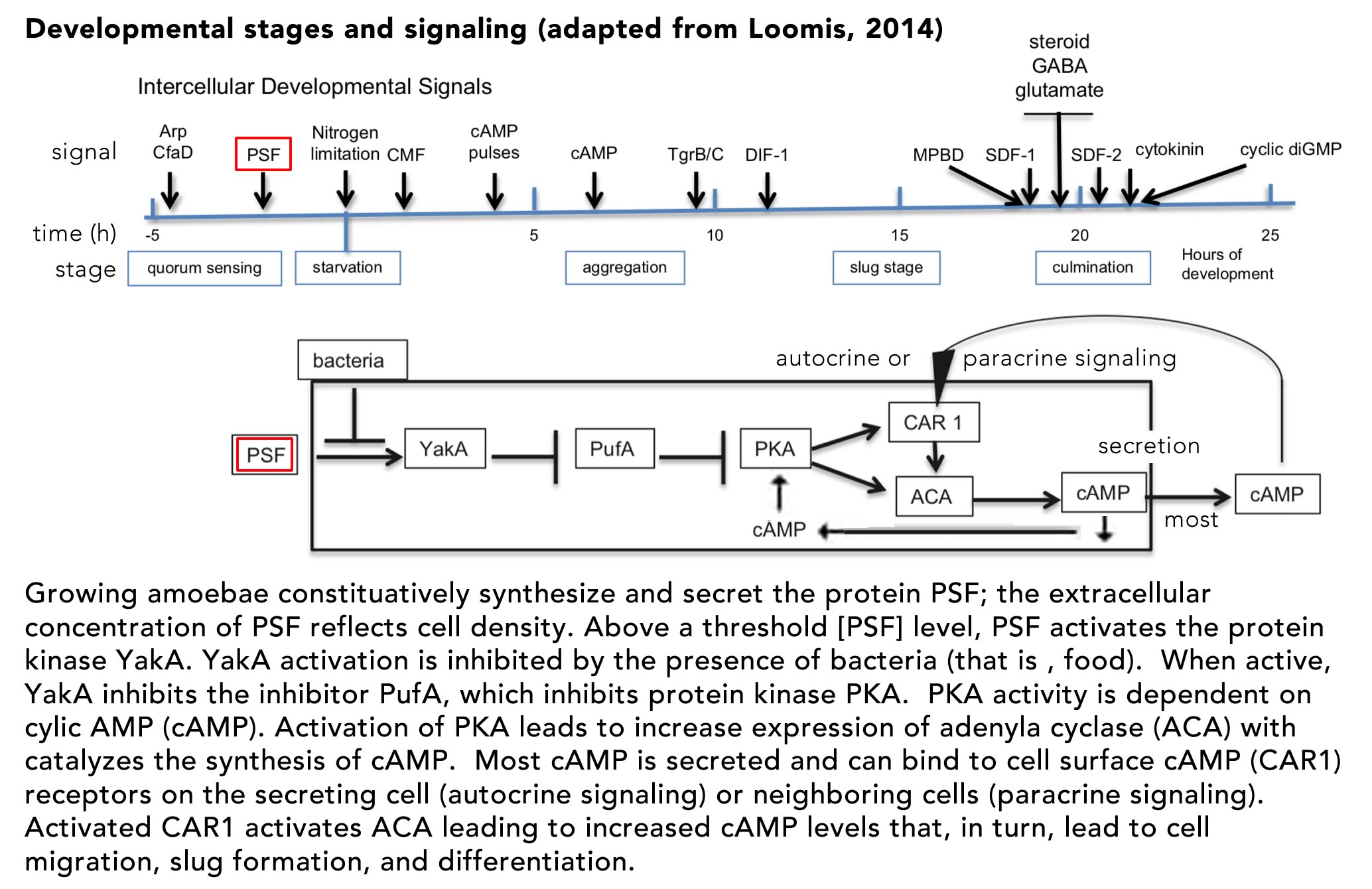
If bacteria, that is food, are present, the activity of PSF is inhibited and cells remain in their single cell state. The key regulator of downstream aggregation and differentiation is the cAMP-dependent protein kinase PKA. In the unicellular state, PKA activity is inhibited by PufA. As PSF increases, while food levels decrease, YakA activity increases, inactivating PufA, leading to increased PKA activity. Active PKA induces the synthesis of two downstream proteins, adenylate cyclase (ACA) and the cAMP receptor (CAR1). ACA catalyzes cAMP synthesis, much of which is secreted from the cell as a signaling molecule. The membrane-bound CAR1 protein acts as a receptor for autocrine (on the cAMP secreting cell) and paracrine (on neighboring cells) signaling. The binding of cAMP to CAR1 leads to further activation of PKA, increasing cAMP synthesis and secretion – a positive feed-back loop. As cAMP levels increase, downstream genes are activated (and inhibited) leading cells to migrate toward one another, their adhesion to form a slug. Once the slug forms and migrates to an appropriate site, the process of differentiation (and death) leading to stalk and spore formation begins. The fates of the aggregated cells is determined stochastically, but social cheaters can arise. Mutations can lead to individuals that avoid becoming stalk cells. In the long run, if all individuals were to become cheaters, it would be impossible to form a stalk, so the purpose of social cooperation would be impossible to achieve. In the face of environmental variation, populations invaded by cheaters are more likely to become extinct. For our purposes the various defenses against cheaters are best left to other courses (see here if interested Strassmann et al., 2000).
Clonal (permanent) multicellularity: The type of multicellularity that most developmental biology courses focus on is what is termed clonal multicellularity – the organism is a clone of an original cell, the zygote, a diploid cell produced by the fusion of sperm and egg, haploid cells formed through the process of meiosis (2). It is during meiosis that most basic genetic processes occur, that is the recombination between maternal and paternal chromosomes leading to the shuffling of alleles along a chromosome, and the independent segregation of chromosomes to form haploid gametes, gametes that are genetically distinct from those present in either parent. Once the zygote forms, subsequent cell divisions involve mitosis, with only a subset of differentiated cells, the cells of the germ line, capable of entering meiosis.
Non-germ line, that is somatic cells, grow and divide. They interact with one another directly and through various signaling processes to produce cells with distinct patterns of gene expression, and so differentiated behaviors. A key difference from a unicellular organism, is that the cells will (largely) stay attached to one another, or to extracellular matrix materials secreted by themselves and their neighbors. The result is ensembles of cells displaying different specializations and behaviors. As such cellular colonies get larger, they face a number of physical constraints – for example, cells are open non-equilibrium systems, to maintain themselves and to grow and reproduce, they need to import matter and energy from the external world. Cells also produce a range of, often toxic, waste products that need to be removed. As the cluster of zygote-derived cells grows larger, and includes more and more cells, some cells will become internal and so cut off from necessary resources. While diffusive processes are often adequate when a cell is bathed in an aqueous solution, they are inadequate for a cell in the interior of a large cell aggregate (3). The limits of diffusive processes necessitate other strategies for resource delivery and waste removal; this includes the formation of tubular vascular systems (such as capillaries, arteries, veins) and contractile systems (hearts and such) to pump fluids through these vessels, as well as cells specialized to process and transport a range of nutrients (such as blood cells). As organisms get larger, their movements require contractile machines (muscle, cartilage, tendons, bones, etc) driving tails, fins, legs, wings, etc. The coordination of such motile systems involves neurons, ganglia, and brains. There is also a need to establish barriers between the insides of an organism and the outside world (skin, pulmonary, and gastrointestinal linings) and the need to protect the interior environment from invading pathogens (the immune system). The process of developing these various systems depends upon controlling patterns of cell growth, division, and specialization (consider the formation of an arm), as well as the controlled elimination of cells (apoptosis), important in morphogenesis (forming fingers from paddle-shaped appendages), the maturation of the immune system (eliminating cells that react against self), and the wiring up, and adaptation of the nervous system. Such changes are analogous to those involved in aggregative multicellularity.
Origins of multicellularity: While aggregative multicellularity involves an extension of quorum sensing and social cooperation between genetically distinct, but related individuals, we can wonder whether similar drivers are responsible for clonal multicellularity. There are a number of imaginable adaptive (evolutionary) drivers but two spring to mind: a way to avoid predators by getting bigger than the predators and as a way to produce varied structures needed to exploit various ecological niches and life styles. An example of the first type of driver of multicellularity is offered by the studies of Boraas et al (1998). They cultured the unicellular green alga Chlorella vulgaris, together with a unicellular predator, the phagotrophic flagellated protist Ochromonas vallescia. After less than 100 generations (cell divisions), they observed the appearance of multicellular, and presumable inedible (or at least less easily edible), forms. Once selected, this trait appears to be stable, such that “colonies retained the eight-celled form indefinitely in continuous culture”. To my knowledge, the genetic basis for this multicellularity remains to be determined.
Cell Differentiation: One feature of simple colonial organisms is that when dissociated into individual cells, each cell is capable of regenerating a new organism. The presence of multiple (closely related) cells in a single colony opens up the possibility of social interactions; this is distinct from the case in aggregative multicellularity, where social cooperation came first. Social cooperation within a clonal metazoan means that most cells “give up” their ability to reproduce a new organism (a process involving meiosis). Such irreversible social interactions mark the transition from a colonial organism to a true multicellular organism. As social integration increases, cells can differentiate so as to perform increasingly specialized functions, functions incompatible with cell division. Think for a moment about a human neuron or skeletal muscle cell – in both cases, cell division is no longer possible (apparently). Nevertheless, the normal functioning of such cells enhances the reproductive success of the organism as a whole – a classic example of inclusive fitness (remember heterocysts?) Modern techniques of single cell sequencing and data analysis have now been employed to map this process of cellular differentiation in increasingly great detail, observations that will inform our later discussions (see Briggs et al., 2018 and future posts). In contrast, the unregulated growth of a cancer cell is an example of an asocial behavior, an asocial behavior that is ultimately futile, except in those rare cases (four known at this point) in which a cancer cell can move from one organism to another (Ujvari et al., 2016).
Unicellular affordances for multicellularity: When considering the design of a developmental biology course, we are faced with the diversity of living organisms – the basic observation that Darwin, Wallace, their progenitors and disciplinary descendants set out to solve. After all there are many millions of different types of organisms; among the multicellular eukaryotes, there are six major group : the ascomycetes and basidiomycetes fungi, the florideophyte red algae, laminarialean brown algae, embryophytic land plants and animals

(Knoll, 2011 ↑). Our focus will be on animals. “All members of Animalia are multicellular, and all are heterotrophs (i.e., they rely directly or indirectly on other organisms for their nourishment). Most ingest food and digest it in an internal cavity.” [Mayer link]. From a macroscopic perspective, most animals have (or had at one time during their development) an anterior to posterior, that is head to tail, axis. Those that can crawl, swim, walk, or fly typically have a dorsal-ventral or back to belly axis, and some have a left-right axis as well.
But to be clear, a discussion of the various types of animals is well beyond the scope of any introductory course in developmental biology, in part because there are 35 (assuming no more are discovered) different “types” (phyla) of animals – nicely illustrated at this website [BBC: 35 types of animals, most of whom are really weird)]. So again, our primary focus will be on one group, the vertebrates – humans are members of this group. We will also consider experimental insights derived from studies of various “model” systems, including organisms from another metazoan group, the ecdysozoa (organisms that shed their outer layer as they grow bigger), a group that includes fruit flies and nematode worms.
My goal will be to ignore most of the specialized terminology found in the scholarly literature, which can rapidly turn a biology course into a vocabulary lesson and that add little to understanding of basic processes relevant to a general understanding of developmental processes (and relevant to human biology, medicine, and biotechnology). This approach is made possible by the discovery that the basic processes associated with animal (and metazoan) development are conserved. In this light, no observation has been more impactful than the discovery that the nature and organization of the genes involved in specifying the head to tail axes of the fruit fly and vertebrates (such as the mouse and human) is extremely similar in terms of genomic organization and function (Lappin et al., 2006 ↓), an observation that we will return to repeatedly. Such molecular similarities extend to cell-cell and cell-matrix adhesion systems, systems that release and respond to various signaling molecules, controlling cell behavior and gene expression, and reflects the evolutionary conservation and the common ancestry of all animals (Brunet and King, 2017; Knoll, 2011).

What can we know about the common ancestor of the animals? Early on in the history of comparative cellular anatomy, the striking structural similarities between the feeding system of choanoflagellate protozoans, a motile (microtubule-based) flagellum a surrounded by a “collar”of microfilament-based microvilli) and a structurally similar organelle in a range of multicellular organisms led to the suggestion that choanoflagellates and animals shared a common ancestor. The advent of genomic sequencing and analysis has only strengthened this hypothesis, namely that choanoflagellates and animals form a unified evolutionary clade, the ‘Choanozoa’ (see tree↑ above)(Brunet and King, 2017). Moreover, “many genes required for animal multicellularity (e.g., tyrosine kinases, cadherins, integrins, and extracellular matrix domains) evolved before animal origins”. The implications is that the Choanozoan ancestor was predisposed to exploit some of the early opportunities offered by clonal multicellularity. These pre-existing affordances, together with newly arising genes and proteins (Long et al., 2013) were exploited in multiple lineages in the generation of multicellular organisms (see Knoll, 2011).
Basically to understand what happened next, some ~600 million years ago or so, we will approach the various processes involved in the shaping of animal development. Because all types of developmental processes, including the unicellular to colonial transition, involve changes in gene expression, we will begin with the factors involved in the regulation of gene expression.
Footnotes:
1). Please excuse the inclusive plural, but it seems appropriate in the context of what I hope will be a highly interactive course.
2). I will explicitly ignore variants as (largely) distractions, better suited for more highly specialized courses.
3). We will return to this problem when (late in the course, I think) we will discuss the properties of induced pluripotent stem cell (iPSC) derived organoids.
Literature cited:
Bonner, J. T. (1998). The origins of multicellularity. Integrative Biology: Issues, News, and Reviews: Published in Association with The Society for Integrative and Comparative Biology 1, 27-36.
Boraas, M. E., Seale, D. B. and Boxhorn, J. E. (1998). Phagotrophy by a flagellate selects for colonial prey: a possible origin of multicellularity. Evolutionary Ecology 12, 153-164.
Briggs, J. A., Weinreb, C., Wagner, D. E., Megason, S., Peshkin, L., Kirschner, M. W. and Klein, A. M. (2018). The dynamics of gene expression in vertebrate embryogenesis at single-cell resolution. Science 360, eaar5780.
Brunet, T. and King, N. (2017). The origin of animal multicellularity and cell differentiation. Developmental cell 43, 124-140.
Gardner, T. S., Cantor, C. R. and Collins, J. J. (2000). Construction of a genetic toggle switch in Escherichia coli. Nature 403, 339-342.
Hillmann, F., Forbes, G., Novohradská, S., Ferling, I., Riege, K., Groth, M., Westermann, M., Marz, M., Spaller, T. and Winckler, T. (2018). Multiple roots of fruiting body formation in Amoebozoa. Genome biology and evolution 10, 591-606.
Knoll, A. H. (2011). The multiple origins of complex multicellularity. Annual Review of Earth and Planetary Sciences 39, 217-239.
Lappin, T. R., Grier, D. G., Thompson, A. and Halliday, H. L. (2006). HOX genes: seductive science, mysterious mechanisms. The Ulster medical journal 75, 23.
Long, M., VanKuren, N. W., Chen, S. and Vibranovski, M. D. (2013). New gene evolution: little did we know. Annual review of genetics 47, 307-333.
Loomis, W. F. (2014). Cell signaling during development of Dictyostelium. Developmental biology 391, 1-16.
Strassmann, J. E., Zhu, Y. and Queller, D. C. (2000). Altruism and social cheating in the social amoeba Dictyostelium discoideum. Nature 408, 965-967.
Ujvari, B., Gatenby, R. A. and Thomas, F. (2016). Transmissible cancers, are they more common than thought? Evolutionary applications 9, 633-634.

3 thoughts on “Aggregative & clonal metazoans: a biofundamentalist perspective”