Insights into student thinking & course design, part of the biofundamentals project.
Something that often eludes both instructors and instructional researchers is a clear appreciation of what it is that students do and do not know, what ideas they can and cannot call upon to solve problems and generate clear, coherent, and plausible explanations. What information – thought to have been presented effectively through past instruction, appears to be unavailable to students. As an example, few instructors would believe that students completing college level chemistry could possibly be confused about the differences between covalent and non-covalent molecular interactions, yet there is good evidence that they are (Williams et al., 2015). Unless these ideas, together with their conceptual bases and practical applications, are explicitly called out in the design and implementation of instructional materials, they often fail to become a working (relevant) part of the students’ conceptual tool-kit.
To identify ideas involved in understanding biological systems, we are using an upper division undergraduate course in developmental biology (blog link) to provide context; this is a final “capstone” junior/senior level course that comes after students have completed multiple required courses in chemistry and biology. Embryonic development integrates a range of molecular level processes, including the control of gene expression, cellular morphology and dynamics, through intrinsic and extrinsic signaling systems.
A key aspect of the course’s design is the use of formative assessment activities delivered through the beSocratic system. These activities generally include parts in which students are asked to draw a graph or diagram. Students are required to complete tasks before the start of each class meeting; their responses are used to inform in-class discussions, a situation akin to reviewing game film and coaching in sports. Analysis of student drawings and comments, carried out in collaboration with Melanie Cooper and her group at Michigan State University, can reveal unexpected aspects of students’ thinking (e.g. Williams et al., 2015). What emerges from this Socratic give and take is an improved appreciation of the qualities of the tasks that engage students (as well as those that do not), and insights into how students analyze specific tasks, what sets of ideas they see as necessary and which necessary ideas they ignore when generating explanatory and predictive models. Most importantly, they can reveal flaws in how necessary ideas are developed. While at an admittedly early stage in the project, here I sketch out some preliminary findings: the first of these deal with steady state concentration and response dynamics.

The ideas of steady state concentration and pathway dynamics were identified by Loertscher et al (2014)as two of five “threshold concepts” in biochemistry and presumably molecular biology as well. Given the non-equilibrium nature of biological systems, we consider the concentration of a particular molecule in a cell in dynamic terms, a function of its rate of synthesis (or importation from the environment) together with its rate of breakdown. On top of this dynamic, the activity of existing molecules can be regulated through various post-translational mechanisms. All of the populations of molecules within a cell or organism have a characteristic steady state concentration with the exception of genomic DNA, which while synthesized is not, in living organisms, degraded, although it is repaired.
In biological systems, molecules are often characterized by their “half life” but this can be confusing, since it is quite different from the way the term is used in physics, where students are likely to first be introduced to it.[1] Echos from physics can imply that a molecule’s half-life is an intrinsic feature of the molecule, rather than of the system in which the molecule finds itself. The equivalent of half-life would be doubling time, but these terms make sense only under specific conditions. In a system in which synthesis has stopped (synthesis rate = 0) the half life is the time it takes for the number of molecules in the system to decrease by 50%, while in the absence of degradation (degradation rate = 0), the doubling time is the time it takes to double the number of molecules in the system. Both degradation and synthesis rates are regulateable and can vary, often dramatically, in response to various stimuli.

In the case of RNA and polypeptide levels, the synthesis rate is determined by many distinct processes, including effective transcription factor concentrations, the signals that activate transcription factors, rates of binding of transcription factors to transcription factor binding sites (which can involve both DNA sequences and other proteins), as well as relevant binding affinities, and the rates associated with the recruitment and activation of DNA-dependent, RNA polymerase. Once activated, the rate of gene specific RNA synthesis will be influenced by the rate of RNA polymerization (nucleotide bases added per second) and the length of the RNA molecules synthesized. In eukaryotes, the newly formed RNA will generally need to have introns removed through interactions with splicing machinery, as well as other post-transcriptional reactions, after which the processed RNA will be transported from the nucleus to the cytoplasm through the nuclear pore complex. In the cytoplasm there are rates associated with the productive interaction of RNAs with the translational machinery (ribosomes and associated factors), and the rate at which polypeptide synthesis occurs (amino acids added per second) together with the length of the polypeptide synthesized (given that things are complicated enough, I will ignore processes such as those associated with the targeting of membrane proteins and codon usage, although these will be included in a new chapter in biofundamentals reasonably soon, I hope). On the degradative side, there are rates associated with interactions with nucleases (that breakdown RNAs) and proteinases (that breakdown polypeptides). These processes are energy requiring; generally driven by reactions coupled to the hydrolysis of adenosine triphosphate (ATP).

That these processes matter is illustrated nicely in work from Harima and colleagues (2014). The system, involved in the segmentation of the anterior region of the presomitic mesoderm, responds to signaling by activating the Hes7 gene, while the Hes7 gene product act to inhibit Hes7 gene expression. The result is an oscillatory response that is “tuned” by the length of the transcribed region (RNA length). This can be demonstrated experimentally by generating mice in which two of the genes three introns (Hes7-3) or all three introns (intron-less) are removed. Removing introns changes the oscillatory behavior of the system (Hes7 mRNA -blue and Hes7 protein – green)(Harima et al., 2013).
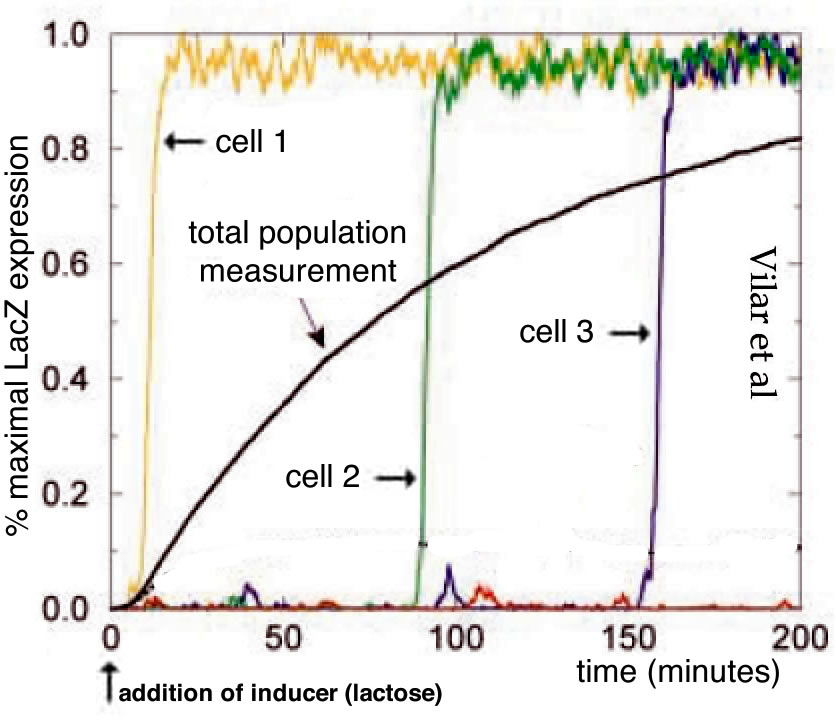
In the context of developmental biology, we use beSocratic activities to ask students to consider a molecule’s steady state concentration as a function of its synthesis and degradation rates, and to predict how the system would change when one or the other is altered. These ideas were presented in the context of observations by Schwanhausser et al (2011) that large discrepancies between steady state RNA and polypeptide concentrations are common and that there is an absence of a correlation between RNA and polypeptide half-lives (we also use these activities to introduce the general idea of correlation). In their responses, it was common to see students’ linking high steady state concentrations exclusively to long half-lives. Ask to consider the implications in terms of system responsiveness (in the specific context of a positively-acting transcription factor and target gene expression), students often presumed that a longer half-life would lead to higher steady state concentration which in turn would lead to increased target gene expression, primarily because collisions between the transcription factor and its DNA-binding sites would increase, leading to higher levels of target gene expression. This is an example of a p-prim (Hammer, 1996) – the heuristic that “more is more”, a presumption that is applicable to many systems.

In biological systems, however, this is generally not the case – responses “saturate”, that is increasing transcription factor concentration (or activity) above a certain level generally does not lead to a proportionate, or any increase in target gene expression. We would not call this a misconception, because this is an example of an idea that is useful in many situations, but generally isn’t in biological systems – where responses are generally inherently limited. The ubiquity and underlying mechanisms of response saturation need to be presented explicitly, and its impact on various processes reinforced repeatedly, preferably by having students use them to solve problems or construct plausible explanations. A related phenomenon that students seemed not to recognize involves the non-linearity of the initial response to a stimulus, in this case, the concentration of transcription factor below which target gene expression is not observed (or it may occur, but only transiently or within a few cells in the population, so as to be undetectable by the techniques used).
So what ideas do students need to call upon when they consider steady state concentration, how it changes, and the impact of such changes on system behavior? It seems we need to go beyond synthesis and degradation rates and include the molecular processes associated with setting the system’s response onset and saturation concentrations. First we need to help students appreciate why such behaviors (onset and saturation) occur – why doesn’t target gene expression begin as soon as a transcription factor appears in a cell? Why does gene expression level off when transcription factor concentrations rise above a certain level? The same questions apply to the types of threshold behaviors often associated with signaling systems. For example, in quorum sensing among unicellular organisms, the response of cells to the signal occurs over a limited concentration range, from off to full on. A related issue is associated with morphogen gradients (concentration gradients over space rather than time), in which there are multiple distinct types of “threshold” responses. One approach might be to develop a model in which we set the onset concentration close to the saturation concentration. The difficulty (or rather instructional challenge) here is that these are often complex processes involving cooperative as well as feedback interactions.
Our initial approach to steady state and thresholds has been to build activities based on the analysis of a regulatory network presented by Saka and Smith (2007), an analysis based on studies of early embryonic development in the frog Xenopus laevis. We chose the system because of its simplicity, involving only four components (although there are many other proteins associated with the actual system). Saka and Smith modeled the regulatory network controlling the expression of the transcription factor proteins Goosecoid (Gsc) and Brachyury (Xbra) in response to the secreted signaling protein activin (↓), a member of

the TGFβ superfamily of secreted signaling proteins (see Li and Elowitz, 2019). The network involves the positive action of Xbra on the gene encoding the transcription factor protein Xom. The system’s behavior depends on the values of various parameters, parameters that include response to activator (Activin), rates of synthesis and the half-lives of Gsc, Xbra, and Xom, and the degrees of regulatory cooperativity and responsiveness.

Depending upon these parameters, the system can produce a range of complex responses. In different regimes (→), increasing concentrations of activin (M) can lead, initially, to increasing, but mutually exclusive, expression of either Xba (B) or Gsc (A) as well as sharp transitions in which expression flips from one to the other, as Activin concentration increases, after which the response saturates. There are also conditions at very low Activin concentration (marked by ↑) in which both Xbra and Gsc are expressed at low levels, a situation that students are asked to explain.
Lessons learned: Based on their responses, captured through beSocratic and revealed during in class discussions, it appears that there is a need to be more explicit (early in the course, and perhaps the curriculum as well) when considering the mechanisms associated with response onset and saturation, in the context of how changes in the concentrations of regulatory factors (through changes in synthesis, turn-over, and activity) impact system responses. This may require a more quantitative approach to molecular dynamics and system behaviors. Here we may run into a problem, the often phobic responses of biology majors (and many faculty) to mathematical analyses. Even the simplest of models, such as that of Saka and Smith, require a consideration of factors generally unfamiliar to students, concepts and skills that may well not be emphasized or mastered in prerequisite courses. The trick is to define realistic, attainable, and non-trivial goals – we are certainly not going to succeed in getting late stage molecular biology students with rudimentary math skills to solve systems of differential equations in a developmental biology course. But perhaps we can build up the instincts needed to appreciate the molecular processes involved in the behavior of systems whose behavior evolves overtime in response to various external signals (which is, of course, pretty much every biological system).
Footnotes:
[1] A similar situation exists in the context of the term “spontaneous” in chemistry and biology. In chemistry spontaneous means thermodynamically favorable, while in standard usage (and generally in biology) spontaneous implies that a reaction is proceeding at a measurable, functionally significant rate. Yet another insight that emerged through discussions with Melanie Cooper.
Mike Klymkowsky
Literature cited
Hammer, D. (1996). Misconceptions or p-prims. How might alternative perspectives of cognitive structure influence instructional perceptions and intentions. Journal of the Learning Sciences 5, 97-127.
Harima, Y., Imayoshi, I., Shimojo, H., Kobayashi, T. and Kageyama, R. (2014). The roles and mechanism of ultradian oscillatory expression of the mouse Hes genes. In Seminars in cell & developmental biology, pp. 85-90: Elsevier.
Harima, Y., Takashima, Y., Ueda, Y., Ohtsuka, T. and Kageyama, R. (2013). Accelerating the tempo of the segmentation clock by reducing the number of introns in the Hes7 gene. Cell Reports 3, 1-7.
Li, P. and Elowitz, M. B. (2019). Communication codes in developmental signaling pathways. Development 146, dev170977.
Loertscher, J., Green, D., Lewis, J. E., Lin, S. and Minderhout, V. (2014). Identification of threshold concepts for biochemistry. CBE—Life Sciences Education 13, 516-528.
Saka, Y. and Smith, J. C. (2007). A mechanism for the sharp transition of morphogen gradient interpretation in Xenopus. BMC Dev Biol 7, 47.
Schwanhäusser, B., Busse, D., Li, N., Dittmar, G., Schuchhardt, J., Wolf, J., Chen, W. and Selbach, M. (2011). Global quantification of mammalian gene expression control. Nature 473, 337.
Williams, L. C., Underwood, S. M., Klymkowsky, M. W. and Cooper, M. M. (2015). Are Noncovalent Interactions an Achilles Heel in Chemistry Education? A Comparison of Instructional Approaches. Journal of Chemical Education 92, 1979–1987.