
![]() Embracing the rationalist and empirically-based perspective of science is not easy. Modern science generates disconcerting ideas that can be difficult to accept and often upsetting to philosophical or religious views of what gives meaning to existence [link]. In the context of biological evolutionary mechanisms, the fact that variation is generated by random (stochastic) events, unpredictable at the level of the individual or within small populations, led to the rejection of Darwinian principles by many working scientists around the turn of the 20th century (see Bowler’s The Eclipse of Darwinism + link). Educational research studies, such as our own “Understanding randomness and its impact on student learning“, reinforce the fact that ideas involving stochastic processes relevant to evolutionary, as well as cellular and molecular, biology, are inherently difficult for people to accept (see also: Why being human makes evolution hard to understand). Yet there is no escape from the science-based conclusion that stochastic events provide the raw material upon which evolutionary mechanisms act, as well as playing a key role in a wide range of molecular and cellular level processes, including the origin of various diseases, particularly cancer [Cancer is partly caused by bad luck](1).
Embracing the rationalist and empirically-based perspective of science is not easy. Modern science generates disconcerting ideas that can be difficult to accept and often upsetting to philosophical or religious views of what gives meaning to existence [link]. In the context of biological evolutionary mechanisms, the fact that variation is generated by random (stochastic) events, unpredictable at the level of the individual or within small populations, led to the rejection of Darwinian principles by many working scientists around the turn of the 20th century (see Bowler’s The Eclipse of Darwinism + link). Educational research studies, such as our own “Understanding randomness and its impact on student learning“, reinforce the fact that ideas involving stochastic processes relevant to evolutionary, as well as cellular and molecular, biology, are inherently difficult for people to accept (see also: Why being human makes evolution hard to understand). Yet there is no escape from the science-based conclusion that stochastic events provide the raw material upon which evolutionary mechanisms act, as well as playing a key role in a wide range of molecular and cellular level processes, including the origin of various diseases, particularly cancer [Cancer is partly caused by bad luck](1).

All of which leaves the critical question, at least for educators, of how to best teach students about evolutionary mechanisms and outcomes. The problem becomes all the more urgent given the anti-science posturing of politicians and public “intellectuals”, on both the right and the left, together with various overt and covert attacks on the integrity of science education, such as a new Florida law that lets “anyone in Florida challenge what’s taught in schools”.
Just to be clear, we are not looking for students to simply “believe” in the role of evolutionary processes in generating the diversity of life on Earth, but rather that they develop an understanding of how such processes work and how they make a wide range of observations scientifically intelligible. Of course the end result, unless you are prepared to abandon science altogether, is that you will find yourself forced to seriously consider the implications of inescapable scientific conclusions, no matter how weird and disconcerting they may be.
There are a number of educational strategies, in part depending upon one’s disciplinary perspective, on how to approach teaching evolutionary processes. Here I consider one, based on my background in cell and molecular biology. Genomicus is a web tool that “enables users to navigate in genomes in several dimensions: linearly along chromosome axes, transversely across different species, and chronologically along evolutionary time.” It is one of a number of web-based resources that make it possible to use the avalanche of DNA (gene and genomic) sequence data being generated by the scientific community. For example, the ExAC/Gnomad Browser enables one to examine genetic variation in over 60,000 unrelated people. Such tools supplement and extend the range of tools accessible through the U.S. National Library of Medicine / NIH / National Center for Biotechnology Information (NCBI) web portal (PubMed).
In the biofundamentals / coreBio course (with an evolving text available here), we originally used the observation that members of our subfamily of primates, the Haplorhini or dry nose primates, are, unlike most mammals, dependent on the presence of vitamin C (ascorbic acid) in their diet. Without vitamin C we develop scurvy, a potentially lethal condition. While there may be positive reasons for vitamin C dependence, in biofundamentals we present this observation in the context of small population size and a forgiving environment. A plausible scenario is that the ancestral Haplorhini population the lost the L-gulonolactone oxidase (GULO) gene (see OMIM) necessary for vitamin C synthesis. The remains of the GULO gene found in humans and other Haplorhini genomes is mutated and non-functional, resulting in our requirement for dietary vitamin C.
How, you might ask, can we be so sure? Because we can transfer a functional mouse GULO gene into human cells; the result is that vitamin C dependent human cells become vitamin C independent (see: Functional rescue of vitamin C synthesis deficiency in human cells). This is yet another experimental result, similar to the ability of bacteria to accurately decode a human insulin gene), that supports the explanatory power of an evolutionary perspective (2).
In an environment in which vitamin C is plentiful in a population’s diet, the mutational loss of the GULO gene would be benign, that is, not strongly selected against. In a small population, the stochastic effects of genetic drift can lead to the loss of genetic variants that are not strongly selected for. More to the point, once a gene’s function has been lost due to mutation, it is unlikely, although not impossible, that a subsequent mutation will lead to the repair of the gene. Why? Because there are many more ways to break a molecular machine, such as the GULO enzyme, but only a few ways to repair it. As the ancestor of the Haplorhini diverged from the ancestor of the vitamin C independent Strepsirrhini (wet-nose) group of primates, an event estimated to have occurred around 65 million years ago, its ancestors had to deal with their dietary dependence on vitamin C either by remaining within their original (vitamin C-rich) environment or by adjusting their diet to include an adequate source of vitamin C.
At this point we can start to use Genomicus to examine the results of evolutionary processes (see a YouTube video on using Genomicus)(3). In Genomicus a gene is indicated by a pointed box ![]() ; for simplicity all genes are drawn as if they are the same size (they are not); different genes get different colors and the direction of the box indicates the direction of RNA synthesis, the first stage of gene expression. Each horizontal line in the diagram below represents a segment of a chromosome from a particular species, while the blue lines to the left represent phylogenic (evolutionary) relationships. If we search for the GULO gene in the mouse, we find it and we discover that its orthologs (closely related genes) are found in a wide range of eukaryotes, that is, organisms whose cells have a nucleus (humans are eukaryotes).
; for simplicity all genes are drawn as if they are the same size (they are not); different genes get different colors and the direction of the box indicates the direction of RNA synthesis, the first stage of gene expression. Each horizontal line in the diagram below represents a segment of a chromosome from a particular species, while the blue lines to the left represent phylogenic (evolutionary) relationships. If we search for the GULO gene in the mouse, we find it and we discover that its orthologs (closely related genes) are found in a wide range of eukaryotes, that is, organisms whose cells have a nucleus (humans are eukaryotes).
We find a version of the GULO gene in single-celled eukaryotes, such as baker’s yeast, that appear to have diverged from other eukaryotes about ~1.500,000,000 years ago (1500 million years ago, abbreviated Mya). Among the mammalian genomes sequenced to date, the genes surrounding the GULO gene are (largely) the same, a situation known as synteny (mammals are estimated to have shared a common ancestor about 184 Mya). Since genes can move around in a genome without necessarily disrupting their normal function(s), a topic for another day, synteny between distinct organisms is assumed to reflect the organization of genes in their common ancestor. The synteny around the GULO gene, and the presence of a GULO gene in yeast and other distantly related organisms, suggests that the ability to synthesize vitamin C is a trait conserved from the earliest eukaryotic ancestors.
Now a careful examination of this map (↑) reveals the absence of humans (Homo sapiens) and other Haplorhini primates – Whoa!!! what gives? The explanation is, it turns out, rather simple. Because of mutation, presumably in their common ancestor, there is no functional GULO gene in Haplorhini primates. But the Haplorhini are related to the rest of the mammals, aren’t they? We can test this assumption (and circumvent the absence of a functional GULO gene) by exploiting synteny – we search for other genes present in the syntenic region (↓). What do we find? We find that this region, with the exception of GULO, is present and conserved in the Haplorhini: the syntenic region around the GULO gene lies on human chromosome 8 (highlighted by the red box); the black box indicates the GULO region in the mouse. Similar syntenic regions are found in the homologous (evolutionarily-related) chromosomes of other Haplorhini primates.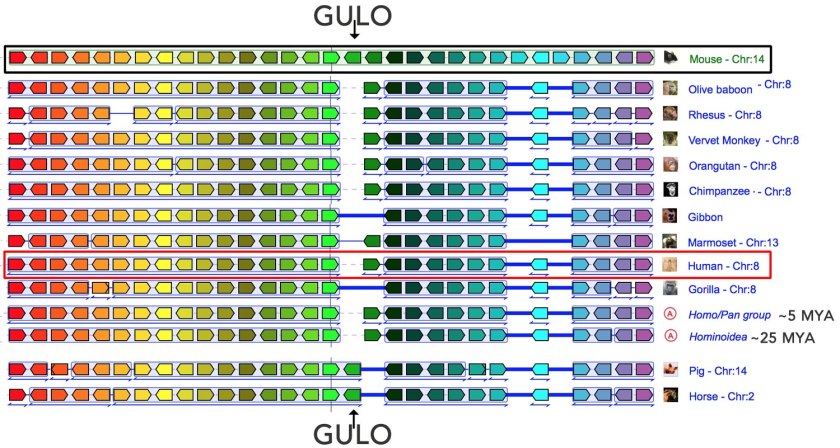
The end result of our Genomicus exercise is a set of molecular level observations, unknown to those who built the original anatomy-based classification scheme, that support the evolutionary relationship between the Haplorhini and more broadly among mammals. Based on these observations, we can make a number of unambiguous and readily testable predictions. A newly discovered Haplorhini primate would be predicted to share a similar syntenic region and to be missing a functional GULO gene, whereas a newly discovered Strepsirrhini primate (or any mammal that does not require dietary ascorbic acid) should have a functional GULO gene within this syntenic region. Similarly, we can explain the genomic similarities between those primates closely related to humans, such as the gorilla, gibbon, orangutan, and chimpanzee, as well as to make testable predictions about the genomic organization of extinct relatives, such as Neanderthals and Denisovians, using DNA recovered from fossils [link].
It remains to be seen how best to use these tools in a classroom context and whether having students use such tools influences their working understanding, and more generally, their acceptance of evolutionary mechanisms. That said, this is an approach that enables students to explore real data and to develop plausible and predictive explanations for a range of genomic discoveries, likely to be relevant both to understanding how humans came to be, and in answering pragmatic questions about the roles of specific mutations and allelic variations in behavior, anatomy, and disease susceptibility.
Some footnotes (figures reinserted 2 November 2020, with minor edits)
(1) Interested in a magnetic bumper image? visit: http://www.cafepress.com/bioliteracy
(2) An insight completely missing (unpredicted and unexplained) by any creationist / intelligent design approach to biology.
(3) Note, I have no connection that I know of with the Genomicus team, but I thank Tyler Square (now at UC Berkeley) for bringing it to my attention.

{kind=link}
An elegant example that demonstrates the importance of understanding evolutionary processes and the scientific method. In the current political climate it is of paramount importance to effectively teach evidence based reasoning. Developing teaching tools that draw upon examples using empirical evidence is of paramount importance as pointed out so clearly above.
LikeLike